
Malattia metabolica del cervello: Attendavamo un documento che spiegasse come l'alluminio sia responsabile delle malattie degenerative del sistema nervoso:. L'articolo è stato pubblicato su ncbi.nlm.nih.gov e Springer Nature Switzerland AG nella sezione Academic Edition. Scritto da: Gerwyn Morris (Tir Na Nog Llanelli UK), Basant K. Puri (Dipartimento di Medicina Imperial College London, Hammersmith Hospital London UK), e Richard E. Frye (College of Medicine, Dipartimento di Pediatria University of Arkansas for Medical Sciences, Arkansas Children's Hospital Research Institute Little Rock USA).
In sintesi: La concettualizzazione del disturbo dello spettro autistico e della malattia di Alzheimer ha subito qualcosa di un cambiamento paradigmatico negli ultimi anni e invece di essere viste come singole malattie con patogenesi unitaria e patofisiologia sono sempre più considerate sindromi eterogenee con una complessa eziopatogenesi multifattoriale, che coinvolge un combinazione complessa e diversificata di fattori genetici, epigenetici e ambientali. Uno di questi fattori ambientali implicati come causa potenziale in entrambe le sindromi è l'alluminio, come elemento o parte di un sale, ricevuto, per esempio, in forma orale o come adiuvante. Tale somministrazione ha il potenziale per indurre patologie attraverso diverse vie come provocare disfunzione e / o attivazione di cellule gliali che svolgono un ruolo indispensabile nella regolazione dell'omeostasi e del neurosviluppo del sistema nervoso centrale. Altre vie includono la generazione di stress ossidativo, l'esaurimento del glutatione ridotto, riduzioni dirette e indirette delle prestazioni e dell'integrità mitocondriale e l'aumento della produzione di citochine proinfiammatorie sia nel cervello che in periferia. Vengono descritti i meccanismi attraverso i quali l'alluminio ambientale potrebbe contribuire allo sviluppo del modello altamente specifico di neuropatologia visto nella malattia di Alzheimer. Inoltre, sono dettagliati diversi meccanismi in base ai quali quantità significative di alluminio introdotte attraverso l'immunizzazione potrebbero produrre neuropatologia cronica in bambini geneticamente suscettibili. Di conseguenza, si raccomanda di interrompere l'uso di sali di alluminio nelle vaccinazioni e che gli adulti debbano adottare misure per ridurre al minimo la loro esposizione all'alluminio ambientale.
introduzione
Disturbo dello spettro autistico (ASD) si riferisce a un gruppo sempre più comune di disturbi eterogenei identificati dalla presenza di menomazioni nelle interazioni sociali e nella comunicazione, insieme a una gamma restrittiva di comportamenti ripetitivi e stereotipati (Zhubi et al 2014 , Ladd-Acosta et al 2014 ). Analisi recenti hanno costantemente dimostrato che la prevalenza di ASD è in aumento. Le stime variano tra uno su 68 (CDC) e uno su 46 (Pelly et al., 2015 ) su uno su 38 (Kim et al., 2011 ) durante periodi di tempo simili. C'è un dibattito sulle ragioni del precipitoso aumento della prevalenza dell'ASD negli ultimi due decenni, con la conclusione che si tratta essenzialmente di un artefatto derivante dallo sviluppo di categorie diagnostiche più ampie e da una maggiore consapevolezza medica (Rutter 2005 ; King and Bearman 2009 ). Tuttavia, analisi condotte da altri gruppi di ricerca hanno suggerito che altri fattori oltre al cambiamento diagnostico possono contribuire a questo aumento della prevalenza (Grether et al., 2009 , Hertz-Picciotto 2009 ). Sebbene le prime stime di ereditarietà per l'ASD fossero elevate (circa il 97%), molti studi genetici completi non sono riusciti a rivelare i geni riproducibili, e finora nessun singolo gene può rappresentare più dell'1% dei casi di ASD, sebbene ci sia una grande quantità di evidenza che le anomalie genetiche giocano un ruolo importante nello sviluppo dei disturbi, il che non sorprende dato che questo è il caso della maggior parte se non di tutte le malattie (Siniscalco et al., 2013 ). Vale anche la pena notare che solo una minoranza di bambini con diagnosi ASD ha una sindrome genetica identificabile (Siniscalco et al., 2013 ) e studi recenti suggeriscono che la resa dei test genetici è bassa anche quando l'analisi dei microarray cromosomici è combinata con l'intero sequenziamento dell'esoma , in particolare nei bambini ASD non-dismorfici (Tammimies et al., 2015 ). Inoltre, alcune anomalie genetiche che aumentano il rischio di sviluppare ASD sono anche fattori di rischio per altre condizioni neurosviluppo o neuroimmune, e sono polimorfismi piuttosto che mutazioni deleteri, essendo presenti anche in individui non affetti (Betancur 2011 , Sahoo et al., 2011 ). Sembrerebbe che questi geni sembrino conferire vulnerabilità a una varietà di disordini dello sviluppo neurologico (Betancur 2011 , Sahoo et al., 2011 ; Rapoport et al., 2012 ). Inoltre, una recente stima di ereditabilità derivante dal più grande studio sui gemelli fino ad oggi ha raggiunto un valore inferiore al 37% (Hallmayer et al., 2011 ), sebbene un altro recente studio abbia calcolato una stima di ereditabilità tra il 40 e il 60% (Klei et al. ).
Il concetto di ASD come malattia di origine puramente genetica ha lasciato il posto all'opinione che, per lo meno, l'eziopatogenesi dell'ASD comporta un'interazione molto complessa tra numerosi geni e fattori di rischio ambientale (Bushnell 2013 ; LaSalle 2013 ). Inoltre, sta diventando sempre più evidente che le alterazioni del paesaggio epigenetico e la disregolazione dei meccanismi epigenetici responsabili dell'espressione genica giocano anche un ruolo importante nell'eziopatogenesi di questi disturbi (Rangasamy et al., 2013 , Flashner et al., 2013 , Siniscalco et al. 2013 ). In uno studio di riferimento su microarray post-mortem, Voineagu e colleghi hanno identificato 444 geni che erano espressi differenzialmente nella corteccia cerebrale e due geni che erano espressi in modo differenziale nel cervelletto, di bambini con ASD rispetto ai controlli neurotipici di età e sesso . Hanno riferito che i pattern di espressione disregolati dei marcatori dei geni immunitari e gliali non erano associati a nessun noto gene di rischio di ASD, quindi i cambiamenti immunitari sono probabilmente fenomeni secondari o il risultato di fattori ambientali (Voineagu et al., 2011 ). In un altro studio, Melnyk e altri hanno esaminato 68 soggetti ASD, 40 fratelli non affetti e 54 controlli di età e sesso, e hanno riportato danni ossidativi al DNA (indicizzati dall'adotto di DNA ossidato 8-oxo-deoxyguanosine) e proteine nei leucociti combinati con ipometilazione globale del DNA specifica per i bambini con ASD (Melnyk et al., 2012 ). È stato anche segnalato il fenomeno dell'ipometilazione globale del DNA nel cervello come fattore di alterazione dell'espressione genica nei bambini con ASD (Ladd-Acosta et al 2014 ; Nardone et al., 2014 ). È anche interessante che quest'ultima squadra abbia riportato l'ipometilazione e la conseguente sovraregolazione dei geni del complemento e del fattore necrotico tumorale alfa (TNF-α), che sono coinvolti nella regolazione dei ruoli dello sviluppo neurologico della microglia e del ridimensionamento sinaptico rispettivamente (Nardone et al. 2014 ).
Ulteriori prove che sottolineano l'importanza delle anormalità epigenetiche nella patogenesi dell'ASD sono state pubblicate da Wong e colleghi ( 2014 ). Questi autori hanno riportato regioni di DNA metilato in modo diverso entro 50 coppie gemellari monozigoti (cioè 100 individui) discordanti per ASD che correlavano con la gravità dei punteggi tratti autistici, sottolineando l'importanza dei fattori epigenetici piuttosto che genetici nella patofisiologia e patogenesi dell'ASD (Wong et al. . 2014 ). Il ruolo della disregolazione epigenetica come fattore importante nella patogenesi dell'ASD è ulteriormente supportato dall'evidenza dell'espressione di microRNA disregolato (miRNA) nei bambini con ASD (Mundalil Vasu et al 2014 ; Mellios and Sur 2012 ). Un recente studio post-mortem ha citato prove di miR-142 ipometilato e sovraregolato nella corteccia frontale (area 10 di Brodmann) nei bambini con ASD, che è interessante in quanto questo miRNA svolge un ruolo importante nella regolazione delle attività dello sviluppo neurologico della microglia e nel mantenerle in uno stato di quiescenza (Mor et al 2015 , Vaishnavi et al., 2013 ; Marrale et al., 2014 ).
Se considerata nel suo insieme, sembra che l'eziologia dell'ASD sia multifattoriale, coinvolgendo una combinazione estremamente complessa e diversificata di fattori genetici, epigenetici, ambientali e immunologici (Flashner et al 2013 , Herbert 2010 , Roberts et al., 2013 ). Nel complesso, la concettualizzazione dell'ASD ha subito un cambiamento di paradigma, e piuttosto che essere vista come una singola malattia con patogenesi unitaria e patofisiologia, è una sindrome clinicamente eterogenea con una complessa eziopatogenesi multifattoriale (Zhubi et al., 2014 Voineagu et al. 2011 ; Betancur 2011 ; Tordjman et al., 2014 ; Ruggeri et al., 2014 ; Georgiades et al., 2013 ). In definitiva, la patologia di un dato bambino probabilmente ha le sue origini in un'interazione dinamica tra un'ampia gamma di diversi agenti ambientali, fattori genetici predisponenti e complessi meccanismi epigenetici come discusso sopra (Zhubi et al., 2014 , Voineagu et al., 2011 , Betancur 2011 ; ; Tordjman et al. 2014 ; Ruggeri et al., 2014 ; Georgiades et al., 2013 ).
Anche la visione dell'ASD come una malattia o malattia che colpisce esclusivamente il cervello sta cambiando. Mentre molti bambini ASD mostrano evidenza di microglia e astrociti attivati, che sono caratteristici di molte malattie neurodegenerative e neurodegenerative (Morgan et al 2012 , Suzuki et al 2013 , Morgan et al 2010 , Morris e Berk 2015 , Morris et al. 2015a ), vi sono anche abbondanti evidenze di anomalie nel sistema immunitario periferico. Tali prove includono dati che dimostrano un'espressione eccessiva di citochine proinfiammatorie (PIC), ridotta espressione di citochine antinfiammatorie, risposte modulate o aumentate di cellule T, risposte di cellule T modificate naturali del killer, risposte del complemento attivato, classe I di maggiore istocompatibilità (MHC) anormalità e aumento degli autoanticorpi sia nella periferia che nel cervello (rivisto in Noriega e Savelkoul 2014 , Careaga e Ashwood 2012 , Estes e McAllister 2015 , Gottfried et al., 2015 ). Lo schema dei polimorfismi a singolo nucleotide (SNP) nei geni immunitari è simile a quelli osservati in diverse malattie autoimmuni come la sclerosi multipla (SM) (Ramos et al., 2012 ). Esistono anche evidenze di attività del recettore di riconoscimento del pattern anormalmente robuste legate alla presenza di SNP nei geni codificanti che portano a risposte immunitarie esagerate (Enstrom et al., 2010 ; Mead and Ashwood 2015 ). È anche degno di nota che i geni che regolano le risposte immunitarie e infiammatorie sono sovraregolati in alcuni bambini con una diagnosi di ASD (Koufaris e Sismani 2015 ) e che la presenza di tali geni espressi in modo anormale può predire lo sviluppo di ASD nei bambini maschi con una precisione dell'83% ( Pramparo et al., 2015 ). È anche interessante che i polimorfismi nei geni citochina e HLA siano associati a risposte insolite ai vaccini (Castiblanco e Anaya 2015 ). L'evidenza di anomalie immunitarie in molti, ma non tutti, i bambini hanno fornito una diagnosi di ASD ha portato alla proposta di un sottotipo neuroimmune di ASD (McDougle et al., 2015 ). Inoltre, diversi ricercatori hanno riportato un'associazione tra insulti infiammatori iniziali e il successivo sviluppo di disturbi immunitari cronici nei bambini con ASD (McDougle et al., 2015 , Gottfried et al., Siniscalco et al., 2013 ). Una possibile spiegazione di questo fenomeno è che una risposta immunitaria insolitamente potente e / o prolungata consente lo sviluppo di danni macromolecolari o tissutali che portano alla formazione di pattern molecolari associati al danno (DAMP) (Lucas et al., 2015 ). La formazione di tali DAMP e la risultante stimolazione cronica dei recettori di riconoscimento del pattern (PRR), che porta allo sviluppo di un "ciclo autotossico" di aumento dell'infiammazione e dello stress ossidativo, è considerata un ruolo importante nel mantenimento e esacerbazione dell'infiammazione sistemica , neuroinfiammazione e neurodegenerazione in una serie di malattie autoimmuni e neurodegenerative come lupus eritematoso sistemico (LES), SM e malattia di Alzheimer (AD) (Venegas and Heneka 2017 , Land 2015 ). Infezioni postnatali frequenti o prolungate sono una fonte ovvia di insulti infiammatori e tali eventi sembrano essere associati a un rischio significativamente maggiore di sviluppare ASD (Abdallah et al., 2012 ; Hadjkacem et al., 2016 ). Tuttavia, altri agenti ambientali potenzialmente associati a un aumentato rischio di sviluppare il fenotipo ASD, come gli organofosfati, il mercurio e l'alluminio, hanno anche la capacità di provocare una risposta immunitaria prolungata o esagerata (Eisenkraft et al., 2013 Kern et al. 2016 ; Shaw e Tomljenovic 2013 ). Sali di alluminio in forma adiuvante funzionanti come DAMP attivano PRR ed esercitano effetti stimolatori profondi sulle risposte immunitarie innate (Powell et al., 2015 ) e quindi potrebbero essere candidati per la generazione di danno tissutale e formazione di DAMP nei bambini con una tendenza sottostante a produrre un esagerato risposta immunitaria. Ci sono anche prove di aumento dei livelli di alluminio nei capelli e nelle urine dei bambini con ASD rispetto ai controlli non affetti (Yasuda e Tsutsui 2013 , Mohamed Fel et al., 2015 Blaurock-Busch et al., Blaurock-Busch et al., 2011 ), anche se questo non è un risultato invariante (Fido e Al-Saad 2005 ; Al-Ayadhi 2005 ). Va inoltre notato che gli adiuvanti di alluminio stanno diventando un fattore scatenante riconosciuto della patologia autoimmune in individui geneticamente predisposti (Morris et al., 2015b ).
L'esposizione intrigante, cronica o cumulativa all'alluminio riflessa da un aumento dei livelli nel liquido cerebrospinale (CSF) e nel siero può essere un fattore ambientale nella patogenesi e patofisiologia della SM, del morbo di Parkinson (PD) e dell'AD (Fulgenzi et al 2014 ; Exley et al. 2006 ; Ahmed e Santosh 2010 ; Yasui et al., 1992 ; Exley and Vickers 2014 ; Basun et al., 1991 ). Esiste una vasta gamma di ricerche che esaminano la potenziale associazione tra una maggiore esposizione all'alluminio ambientale e lo sviluppo dell'ultima di queste malattie. In effetti, una recente meta-analisi che ha coinvolto otto studi di coorte e casi controllati condotti prima del 2015 e che hanno coinvolto 10.567 partecipanti ha concluso che l'aumento dell'alluminio aumentava il rischio di sviluppare l'AD di circa il 71% (Wang et al., 2016 ). Questo sembra un risultato degno di nota alla luce delle prove che indicano che l'AD è anche una sindrome etiologicamente eterogenea (Lam et al., 2013 , Morris e Berk 2015 ), come nel caso del PD (Klein e Lohmann 2009 ; Korczyn e Hassin-Baer 2015 ) e MS (Paz Soldan e Rodriguez 2002 ). Quindi stabilire un'associazione tra AD e un singolo fattore ambientale in uno studio trasversale quando una molteplicità di tali fattori può essere coinvolta in diversi pazienti è un'impresa difficile. Gran parte delle prove in vivo che esaminano i meccanismi alla base degli effetti patologici dell'esposizione all'alluminio sono state ottenute nell'area della ricerca umana e animale nella patogenesi dell'AD. Tale evidenza include l'induzione dello stress ossidativo, la disfunzione mitocondriale, l'attivazione microgliale e la disregolazione funzionale della microglia (Morris e Berk 2016 ). Questo può essere molto rilevante in quanto molti bambini con ASD manifestano anche stress ossidativo (rivisto in Depino 2013 , Rossignol e Frye 2014 , Frustaci et al., 2012 ), disfunzione mitocondriale (recensione di Goh et al 2014 , Chen et al., 2015 ; Guevara -Campos et al., 2013 ) e attività microgliale anormale come discusso sopra.
In questo articolo miriamo a rivedere le prove disponibili per stabilire un'associazione tra l'aumento dell'esposizione all'alluminio e un aumentato rischio di sviluppare l'AD e le prove volte a chiarire i potenziali meccanismi patofisiologici mediante i quali l'alluminio potrebbe essere un elemento nello sviluppo della malattia in almeno alcune persone L'obiettivo di questa parte del documento è di informare i lettori interessati alla patogenesi e alla patofisiologia dell'ASD che potrebbero non essere consapevoli delle preoccupazioni relative all'alluminio nella patogenesi di condizioni diverse dall'ASD. Miriamo anche ad evidenziare l'accumulo di prove che suggeriscono che gli adiuvanti di alluminio possono scatenare una grave patologia autoimmune o autoinfiammatoria nelle persone geneticamente suscettibili, che è una crescente area di preoccupazione. Passeremo quindi a considerare le prove che suggeriscono un'associazione tra l'aumento dell'uso di adiuvanti di sale di alluminio e un'aumentata incidenza di ASD prima di passare a toccare brevemente la sicurezza o meno dei vaccini in persone con predisposizione all'autoimmunità e una gamma di polimorfismi nei geni immunitari. Ciò sembrerebbe appropriato alla luce dei cambiamenti nella concettualizzazione dell'ASD come una sindrome con una molteplicità di cause potenziali e una crescente conoscenza degli effetti della variazione genetica nel sistema immunitario e della risposta ai vaccini. Il resto del documento si concentrerà sui meccanismi attraverso i quali l'aumento dell'esposizione all'alluminio potrebbe essere un fattore scatenante ambientale dell'ASD in almeno alcuni bambini con una serie di anomalie nelle prestazioni del loro sistema immunitario.
Sezione 1. Prove che evidenziano le proprietà neurotossiche dell'alluminio
Prove di un'associazione tra esposizione cronica dell'alluminio e sviluppo di AD
L'alluminio metallico del blocco metallico, che è il terzo elemento più frequente nella crosta terrestre, si trova naturalmente nella bauxite minerale, varie argille e minerali alumino-silicati e ha uno stato di ossidazione preferito di +3. Diversi autori hanno riportato una forte correlazione positiva tra il livello di alluminio nell'acqua potabile e l'incidenza di AD in tutto il mondo, tra cui Regno Unito, Canada, Norvegia e Francia (Flaten 2001 , Kawahara e Kato-Negishi 2011 ). L'associazione più recentemente segnalata è stata pubblicata da Rondeau et al. ( 2009 ), che ha dimostrato che l'elevato consumo giornaliero di alluminio nell'acqua potabile era associato a un rischio significativamente maggiore di sviluppare un danno cognitivo lieve o AD in uno studio longitudinale di coorte francese di 15 anni che coinvolgeva 1925 reclute (Rondeau et al., 2009 ).
Esistono notevoli prove in vitro e in vivo che dimostrano che gli ioni di alluminio inibiscono la defosforilazione del tau, potenziano lo sviluppo di grovigli neurofibrillari (NFT), causano l'accumulo di proteina beta amiloide e accelerano la formazione di placche amiloidi (Kawahara 2005 ; Exley 2005 ) . Nonostante tali prove, l'entusiasmo per l'alluminio come fattore nella patogenesi della DA diminuì in gran parte basandosi su dati che suggerivano che i livelli di alluminio non erano più alti nel cervello dei pazienti affetti da AD rispetto ai controlli sani e l'incapacità di rilevare l'alluminio nelle NFT e nelle placche amiloidi nel tessuto post-mortem a livelli più alti nell'AD che in controlli per età e sesso (Yumoto et al., 2009 ). In particolare, nello studio di Jacobs et al. Non è stata trovata alcuna evidenza di aumento dei livelli di alluminio nel cervello in AD, utilizzando spettrofotometria di assorbimento atomico senza fiamma. ( 1989 ). Inoltre, alti livelli di alluminio nei nuclei delle placche amiloidi o neuritici (senili) non sono stati riportati in diversi studi che impiegano in vario modo l'analisi microscopia a scansione protonica (Lovell et al., 1993 ), analisi microsonda a dispersione di energia a dispersione (Jacobs et al. 1989 ), analisi con microscopia elettronica (Chafi et al., 1991 ) o microscopia nucleare mediante emissione di raggi X indotta da particelle, spettrometria di retrodiffusione di Rutherford e microscopia a scansione ionica a trasmissione (Landsberg et al., 1992 ). Al contrario, l'aumento dell'alluminio della placca è stato riportato in AD utilizzando un sistema microanalitico a dispersione di energia a raggi X (Edwardson et al., 1986 ) e un metodo basato sulla spettrometria di massa al plasma accoppiata induttivamente combinata con iniezione di flusso (Beauchemin e Kisilevsky 1998 ). Allo stesso modo, i risultati relativi all'aumento dell'alluminio nelle NFT sono incoerenti, con una scoperta positiva che utilizza l'analisi di massa con microsonda laser (Good et al., 1992 ), risultati negativi con analisi di microsonda elettronica e microsonda ionica (Chafi et al., 1991 ) e un intermedio ritrovamento (cioè lieve aumento) di nuovo usando l'analisi di massa con microsonda laser (Lovell et al., 1993 ). Uno studio istochimico dei neuroni dell'ippocampo AD ha riportato evidenza di alluminio nei nucleoli e nelle NFT (Walton 2006 ).
Nonostante i risultati di cui sopra, alcuni recenti studi di ricerca che utilizzano tecniche più sensibili hanno rilevato l'alluminio nel cervello dei pazienti affetti da AD all'interno di placche, NFT e altrove a livelli significativamente più elevati rispetto ai controlli non affetti corrispondenti a età e sesso (Yumoto et al., 2009 ; Bouras et al., 1997 ). Inoltre, una serie di studi che riportano gli effetti dell'esposizione dell'alluminio negli animali hanno dimostrato lo sviluppo di patologie AD e Alzheimer nei roditori (Al-Olayan et al., 2015 ; Abd-Elghaffar et al., 2005 ; Sumathi et al., 2015 ; Lu et al., 2014 ; Exley and Vickers 2014 ; Exley and Esiri 2006 ). Studi su animali hanno anche rivelato che l'alluminio somministrato per via orale o tramite iniezione ha ridotto significativamente i livelli ridotti di glutatione e le attività di catalasi, superossido dismutasi, glutatione perossidasi e glutatione reduttasi e ha aumentato i livelli di ossido nitrico (NO), PIC e perossidazione lipidica (Sumathi et Al. 2015 ; Al-Olayan et al., 2015 ). Inoltre, l'esame istologico ha rivelato l'apoptosi dei neuroni ippocampali e cerebrali corticali e la presenza di NFT, deposizione di placche amiloidi, degenerazione delle cellule di Schwann e demielinizzazione delle fibre nervose (Abd-Elghaffar et al., 2005 ).
La conoscenza riguardante i possibili meccanismi attraverso i quali l'esposizione dell'alluminio potrebbe provocare alcune delle caratteristiche caratteristiche alla base della fisiopatologia dell'AD si è anche evoluta. In un recente articolo, Zhao et al. ( 2014 ) hanno riportato sulla capacità di livelli fisiologicamente realistici di alluminio di provocare l'aggregazione di monomeri di Ap42 in strutture dimeriche, oligomeriche e, in definitiva, fibrillari. Questo gruppo di autori ha anche citato la diminuzione dell'espressione del recettore innescante espresso nelle cellule mieloidi / microgliali-2 (TREM2) nella microglia in seguito alla sovraregolazione del miR-34a come meccanismo alla base della compromissione della clearance mediata dalla microglia del peptide Aβ42 dal cervello causata dal prolungato esposizione all'alluminio a concentrazioni nanomolari (Zhao et al., 2014 ).
Studi umani in vivo hanno anche riportato anomalie specifiche correlate all'alluminio nel cervello dei pazienti con AD. L'associazione tra esposizione prolungata all'alluminio ambientale e aumento dei livelli di sottospecie tau fosforilate nei linfociti del sangue è stata anche riportata in un recente studio che ha coinvolto 66 lavoratori dell'alluminio in pensione (Lu et al., 2014 ). Inoltre, è stato dimostrato che la ferritina nel plasma dei pazienti con AD, in particolare quelli con AD lieve, contiene concentrazioni di alluminio significativamente più elevate rispetto alla ferritina plasmatica derivante da controlli di età e sesso che, dato il ruolo chiave di questa proteina nella regolazione dell'omeostasi del metallo, può essere una scoperta cruciale; il riscontro di un livello più elevato nell'AD lieve rispetto all'AD severa può anche indicare una prima fase in cui si verifica un sovraccarico di ferritina di alluminio, seguito da una fase in cui la ferritina con capacità funzionale ridotta rilascia alluminio (De Sole et al., 2013) ). È interessante notare che la capacità dell'alluminio di interrompere l'attività di ferritina e transferrina, con la conseguente rottura dell'omeostasi del ferro, è stata dimostrata in una serie di studi che hanno implicato l'alluminio come potenziale agente causale in alcuni tipi di cellule di cancro al seno e in primarie carcinomi mammari invasivi e carcinoma duttale in situ (Darbre et al., 2013 ; Darbre et al., 2011 ; Mannello et al., 2013 ).
Va anche notato che, fino a poco tempo fa, mancava una spiegazione che potesse spiegare gli effetti selettivi dell'AD in varie regioni del cervello. Tuttavia, in uno studio che utilizza la spettroscopia di assorbimento atomico elettrotermico del contenuto di alluminio delle pareti arteriose di otto arterie che forniscono il cervello, si è riscontrato che la concentrazione di alluminio è molto più alta nell'arteria cerebrale posteriore (arteria cerebri posteriore), che fornisce l'ippocampo , nei pazienti con AD in stadio avanzato rispetto ai controlli per età e sesso (Bhattacharjee et al., 2013 ). Questo studio è particolarmente interessante perché, considerati nel loro complesso, i dati indicano la presenza di meccanismi biochimici nelle cellule endoteliali che forniscono la vascolarizzazione cerebrale che consente il legame dell'alluminio con aree selezionate come l'ippocampo, noto per svolgere un ruolo importante in la patogenesi della malattia (Bhattacharjee et al., 2013 ).
Non c'è dubbio che il peso delle prove che implicano l'alluminio nella causalità di AD in almeno alcuni pazienti sia in aumento. Tuttavia, al momento attuale, nonostante un'analisi che utilizzava i criteri di causalità di Hill per concludere che l'alluminio svolgesse un ruolo causale nello sviluppo di AD (Walton 2014 ), al momento non esiste un consenso universale sull'argomento e sembra ragionevole concludere che vi sia un legame correlativo tra alluminio e AD, ma che attualmente questa associazione non costituisce una relazione causale. Esiste tuttavia un accumulo di prove che suggeriscono che l'alluminio in forma adiuvante può provocare condizioni autoimmuni sistematiche e sintomatiche in individui geneticamente predisposti e discuteremo ora di questo fenomeno.
Il coinvolgimento di adiuvanti di alluminio nello sviluppo della sindrome autoimmune indotta da adiuvanti (ASIA)
Prove che dimostrano lo sviluppo di condizioni autoimmuni o autoinfiammatorie croniche in seguito all'esposizione ambientale ai sali di alluminio, e anzi ad altri adiuvanti, stanno diventando sempre più motivo di preoccupazione (Zafrir et al., 2012 ; Cerpa-Cruz et al., 2013 Jensen-Jarolim 2015 ; Willhite et al., 2014 ). Molte di queste evidenze sono state presentate nel contesto della "sindrome autoimmune (autoinfiammatoria) indotta dagli adiuvanti" (ASIA), che comprende un ampio spettro di malattie immuno-mediate innescate dall'esposizione a coadiuvanti medici, cosmetici o ambientali come sali di alluminio, composti di silicio o muffe da interno (Agmon-Levin et al., 2009 ). L'ASIA è caratterizzata da manifestazioni specifiche e non specifiche di malattie autoimmuni come stanchezza cronica, mialgia, artralgie, compromissione neurocognitiva, sintomi respiratori, sintomi gastrointestinali, segni dermatologici e sviluppo di autoanticorpi (israeliani 2012 ).
Le condizioni mediche considerate da alcuni come parte della sindrome includono fenomeni di post-vaccinazione, sindrome della Guerra del Golfo, miofascite macrofagica, sindrome antifosfolipidica, siliconosi e possibile sindrome da affaticamento cronico (encefalomielite mialgica) e sindrome fibromialgica (Cruz-Tapias et al., 2013 ; -Lastra et al., 2013 ). È interessante notare che i dati di modelli animali suggeriscono che gli adiuvanti possono giocare un ruolo nello sviluppo di malattie autoimmuni sindromiche come il SLE, la sindrome di Sjögren e l'artrite reumatoide in alcuni pazienti (Cruz-Tapias et al., 2013 ; Bagavant et al. 2014 ).
Un tempo si pensava che gli adiuvanti rappresentassero una minima o nessuna minaccia indipendente come fattori guida della patologia. Sfortunatamente, studi di modelli animali e umani hanno dimostrato la capacità di alcuni di loro di indurre autoimmunità e malattie immuno-mediate (Agmon-Levin et al., 2009 ; Elkayam et al., 2011 ). I meccanismi alla base dell'immunotossicità indotta da adiuvante sembrano essere piuttosto vari, ma chiaramente influenzano sia le risposte immunitarie innate che quelle umorali (Marrack et al., 2009 ; Kool et al., 2008 ; Eisenbarth et al., 2008 ). Va detto tuttavia che l'esposizione adiuvante di per sé non sembra causare problemi per la stragrande maggioranza delle persone e lo sviluppo di ASIA sembra dipendere dalla predisposizione genetica o da co-fattori ambientali ancora non determinati (Perricone et al., 2013 ; Esposito et al 2014 ; Shoenfeld e Agmon-Levin 2011 ; Soriano et al., 2015 ).
Diversi autori hanno esaminato pazienti con diagnosi di malattie autoimmuni o altre malattie immuno-mediate dopo l'immunizzazione del virus dell'epatite B (Zafrir et al., 2012 ; Agmon-Levin et al., 2009 ; Agmon-Levin et al., 2014 ). Il più grande studio di questo tipo ha valutato le cartelle cliniche di 93 pazienti e riportato i tassi di prevalenza di diverse manifestazioni come segue: neurologico 67%; sintomi generali 60%; muscoloscheletrico 60%; gastrointestinale 51%; affaticamento 42%, oftalmologico 32%; muco-cutaneo 30%; disturbo del sonno 19%; psichiatrico 16%; e reazioni locali dell'11% (Zafrir et al., 2012 ). Titoli di autoanticorpi elevati sono stati documentati anche nei sieri nell'80% dei pazienti. Diversi adiuvanti del vaccino sono stati anche implicati nello sviluppo di malattie autoimmuni che si trovano al di fuori dell'ASIA, in particolare l'ASD che è un argomento che ora consideriamo.
Coadiuvanti di alluminio nella patogenesi dell'ASD
Dal punto di vista degli adiuvanti di alluminio come potenziale fattore contributivo nello sviluppo dell'ASD (Shaw e Tomljenovic 2013 ), un'analisi recente che applica i criteri di Hill per stabilire la causalità ha riportato che i bambini che vivono in paesi con la più alta prevalenza di ASD sembrano avere la maggiore esposizione alluminio a base di vaccino. Forse ancora più importante, l'aumento dell'esposizione ad adiuvanti di alluminio ha mostrato una significativa correlazione positiva con l'aumento della prevalenza di ASD negli USA registrato negli ultimi 20 anni ( r = 0,92, p <0,0001). Un'analisi più ampia ha rivelato la presenza di correlazione positiva significativa tra i livelli di alluminio in forma adiuvante somministrato ai bambini in età prescolare a circa 3-4 mesi e la prevalenza di ASD esistente in sette principali paesi occidentali ( r = 0,89-0,94, p = 0,0018 0,0248) (Shaw e Tomljenovic 2013 ).
Taylor e colleghi, in una meta-analisi di 10 studi pre-selezionati prevalentemente retrospettivi, non hanno riportato alcuna relazione causale tra una serie di vaccini contenenti mercurio e una serie di condizioni dello sviluppo neurologico come il ritardo dello sviluppo pervasivo (PDD), disturbo da deficit di attenzione / iperattività ( ADHD), disturbo autistico e ASD come diagnosticato con diversi criteri (Taylor et al., 2014 ).
Vale la pena notare che le domande poste dai due gruppi precedenti sono diverse, con il primo che si concentra interamente sull'autismo usando i criteri diagnostici correnti, mentre il secondo ha usato una gamma di diverse definizioni di casi di autismo e condizioni simili all'autismo e includeva anche bambini con PDD. . In effetti, un ampio studio incluso nell'analisi si concentrava interamente sulla PDD (Smeeth et al., 2004 ), mentre altri due si concentravano sulle condizioni generali dello sviluppo neurologico (Andrews et al., 2004 ; Verstraeten et al., 2003 ). È anche giusto dire che le conclusioni di un altro studio (DeStefano 2007 ) sono state messe in discussione e una nuova analisi dei dati ha rivelato un'associazione significativa tra una prima immunizzazione del morbillo, della parotite e della rosolia (MMR) prima dei 36 mesi di età in maschi afro-americani e una diagnosi di ASD (Hooker 2014 ), anche se va notato che quest'ultimo studio è stato successivamente ritirato. Sembrerebbe che gli statistici coinvolti nella meta-analisi di Taylor et al. ( 2014 ) hanno visto PDD, ADHD e ASD come essenzialmente la stessa condizione, o, l'interpretazione di questo e altri dati si basa sul concetto di ASD come entità di malattia discreta che viene sempre più chiamata in causa (Zhubi et al., 2014 ; Voineagu 2012 ; Betancur 2011 ; Tordjman et al. 2014 ; Ruggeri et al., 2014 ; Georgiades et al., 2013 ). Tuttavia, nonostante questi problemi coloro che credono che gli adiuvanti causino "autismo" hanno una domanda che rimane senza risposta, e cioè che se l'alluminio o altri adiuvanti di per sé provocano il sistema nervoso centrale e la patologia periferica, perché la prevalenza dell'ASD non è lontana più in alto di adesso? Data l'ubiquità della vaccinazione ci si aspetterebbe che quasi tutti i bambini ne sarebbero colpiti. Questo fatto da solo significa che è altamente improbabile che gli adiuvanti siano la causa principale dell'ASD. Tuttavia, la ricerca all'interno del paradigma ASIA e oltre suggerisce che gli adiuvanti di alluminio, e di fatto la vaccinazione di per sé , possono causare gravi patologie a lungo termine in persone con una certa vulnerabilità genetica, specialmente nel caso di malattie autoimmuni latenti o subcliniche, e passiamo ora brevemente al dettaglio di tali prove.
Sezione 2. Effetti patologici dei vaccini in persone con predisposizione all'autoimmunità
Langer-Gould e colleghi hanno esaminato le cartelle cliniche di 780 pazienti con SM di nuova diagnosi, sindrome clinicamente isolata (neurite ottica, mielite trasversa e sindrome monofocale o multifocale clinicamente isolata) o encefalomielite acuta disseminata (ADEM) e hanno concluso che i vaccini possono accelerare o precipitare la transizione tra condizioni autoimmuni sintomatiche subcliniche e conclamate entro i primi 30 giorni post-immunizzazione, in particolare in quelli di età inferiore ai 50 anni (Langer-Gould et al., 2014 ). Diversi altri autori hanno riportato un'associazione tra il vaccino quadrivalente del papilloma umano e lo sviluppo di diverse malattie autoimmuni, tra cui il morbo di Raynaud, la sindrome di Behçet, il diabete mellito di tipo 1 e la sindrome di Hashimoto (Arnheim-Dahlstrom et al., 2013 ; Chao et al., 2012 ). Tuttavia, ancora una volta sembrerebbe che i pazienti colpiti mostrassero segni di autoimmunità subclinica prima della vaccinazione che possono avere successivamente attivato la malattia attiva (Chao et al., 2012 ; Arnheim-Dahlstrom et al., 2013 ). Molti gruppi di ricerca che hanno esaminato i dati sugli eventi avversi hanno raggiunto conclusioni simili (Pellegrino et al., 2015 , Petrovsky 2015 , Guimaraes et al., 2015 ). Grimaldi-Bensouda e colleghi hanno trovato un'associazione positiva tra una storia personale e familiare di malattie autoimmuni e lo sviluppo di diverse malattie autoimmuni post-vaccinazione (Grimaldi-Bensouda et al., 2014 ). È interessante notare che, forse in modo rassicurante, uno studio prospettico longitudinale controllato che esamina pazienti inizialmente non affetti senza evidenza di malattia autoimmune conclamata o nascosta non ha dimostrato alcuna associazione tra la vaccinazione e lo sviluppo di ADEM o altre condizioni autoimmuni (Scheller et al., 2015 ). Tuttavia, come notato in precedenza, vi sono prove considerevoli che i vaccini, o più probabilmente adiuvanti del vaccino, possono scatenare sequele autoimmuni specifiche in persone geneticamente o epigeneticamente vulnerabili (Pellegrino et al., 2015 , Petrovsky 2015 , Guimaraes et al., 2015 ). Discuteremo ora sui possibili meccanismi che potrebbero sostenere questo effetto.
Polimorfismi nell'antigene dei leucociti umani (HLA) e recettore Toll-like (TLR) e risposta immunitaria alla vaccinazione
Esiste una vasta serie di dati che dimostrano che le risposte immunitarie e infiammatorie ai vaccini come l'MMR sono pesantemente influenzate da polimorfismi nella regione HLA e in geni che codificano proteine effettrici come citochine e PRR che hanno la capacità di riconoscere e attivarsi da agenti patogeni conservati modelli molecolari associati (PAMP) per produrre molecole di risposta immunitaria come PIC e interferoni (Haralambieva et al 2013 , Lucas and Maes 2013 ). Esempi di PRR includono recettori legati alla membrana come TLR (es. TLR-4) e recettori citosolici come recettori del gene retinoico inducibile all'acido (RIG) (Kumar et al., 2013 ). I lettori interessati ad esaminare le prove che pretendono di dimostrare un'associazione tra i polimorfismi HLA e la risposta insolita al vaccino MMR si riferiscono ad un'eccellente revisione di Castiblanco e Anaya 2015 . Ricerche approfondite hanno anche rivelato che le risposte immunitarie ai vaccini di per sé in un dato individuo sono determinate da polimorfismi e schemi di metilazione nella regione HLA, citochine e geni TLR accoppiati con la composizione del microbioma, la presenza di co-infezioni e un intero host di variabili ambientali (recensione Poland et al., 2013 ). Queste osservazioni forniscono la base per un meccanismo in base al quale gli adiuvanti potrebbero provocare una risposta anormale nelle persone con determinati polimorfismi e / o schemi di metilazione nella regione HLA, citochine e TLR che portano a un'immunità immunitaria eccessivamente potente e / o prolungata con conseguente danno tissutale e generazione di DAMPs, come S100b, con conseguente sviluppo di patologie croniche immunitarie e infiammatorie (Lucas and Maes 2013 , Lucas et al., 2015 ). Questo sarà il tema sviluppato nel resto di questo articolo. In primo luogo, tuttavia, è opportuno considerare se l'alluminio in forma ambientale o adiuvante possiede effettivamente la capacità di generare la gamma di patologie osservate in alcuni bambini con una diagnosi ASD, che ora considereremo.
Sezione 3. Esposizione cronica dell'alluminio e sviluppo dello stress ossidativo cronico, disfunzione mitocondriale e gliopatologia
Esposizione dell'alluminio che provoca produzione di PIC e chemochine
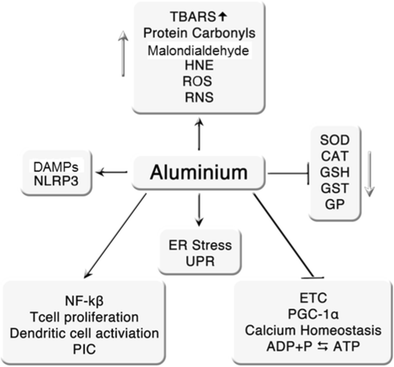
Gli adiuvanti contenenti sali in alluminio inducono la produzione di interleuchine (IL) IL-1β, IL-8 e IL-18 in cellule dendritiche e macrofagi stimolati con TLR (Kuroda et al 2011 , Li et al., 2008 , Sharp et al. 2009 ). Ora ci sono dati considerevoli, anche se in vitro , che dimostrano che tale attivazione dipende dall'attivazione dell'inflammasoma del dominio del recettore pirinico del dominio-3 (NLRP3) di oligomerizzazione nucleotidico (Kool et al. 2008a ; Franchi and Nunez 2008 ). I sali di alluminio possono attivare l'inflammasoma NLRP3 attraverso un numero di percorsi diversi. Questi includono destabilizzazione dei fagosomi, acidificazione dei lisosomi e aumento dei livelli delle specie reattive dell'ossigeno (ROS) (Kool et al., 2012 ; Hornung et al., 2008 ; Sharp et al., 2009 ). In vivo , l'idrossido di alluminio sembra indurre l'attivazione delle cellule dendritiche e delle cellule T almeno in parte tramite l'attivazione di NLRP3 (Kool et al. 2008a Eisenbarth et al., 2008 ) sebbene percorsi alternativi come il motivo di attivazione a base di tirosina (ITAM) e l'interferone degli immunorecettori l'attivazione del fattore di risposta 3 (irf3) sembra essere coinvolta (Kuroda et al., 2011 ; Marichal et al., 2011 ). Gli adiuvanti di alluminio provocano anche una risposta immunitaria attraverso la generazione di DAMP, in particolare acido urico e DNA ospite (al di fuori dei nuclei cellulari e dei mitocondri) (Kool et al., 2012 ; Kool et al., 2008b ). Esiste un insieme di prove che dimostrano che l'acido urico e il DNA sono rilasciati in vivo dopo l'iniezione di idrossido di alluminio (Marichal et al., 2011 ; Kool et al., 2008b ). L'acido urico è un DAMP sintetizzato durante il catabolismo del nucleotide purinico la cui concentrazione aumenta durante lo stress cellulare, come nel sito di iniezione (Kool et al., 2008b ). La rilevanza funzionale dei livelli di acido urico nell'innalzamento del priming delle cellule T e nell'istigazione delle risposte immunitarie umorali è stata ripetutamente dimostrata (Kool et al., 2008b ; Munks et al., 2010 ). Sia l'acido urico che l'idrossido di alluminio possono attivare autonomamente l'inflammasoma NLRP3, inducendo la secrezione di IL-1β (Kool et al. 2008a ; Franchi and Nunez 2008 ). I cristalli di acido urico somministrati sotto forma di adiuvante possono anche indurre risposte di complemento e un modello di differenziazione cellulare di tipo T helper di tipo 2 (T h 2) (Kool et al., 2011 , Shi et al., 2003 ). Il DNA ospite rilasciato nello spazio intracellulare in seguito alla necrosi cellulare agisce anche come DAMP. Testimonianze della natura altamente immunogenica del DNA a doppio filamento sono dimostrate dai dati che dimostrano che può essere usato come sostituto dell'idrossido di alluminio come coadiuvante del vaccino (Marichal et al., 2011 ). Il DNA citosolico a doppio filamento è rilevato da un numero di PRR tra cui TLR-9, che porta alla produzione di PIC attraverso l'attivazione del fattore nucleare potenziatore della catena kappa-luce delle cellule B attivate (NF-κB) o dell'interferone-beta ( IFN-β) tramite l'attivazione di irf3 (Stetson e Medzhitov 2006 ; Thompson et al., 2011 ).
Esposizione dell'alluminio e generazione dello stress ossidativo
Il danno ossidativo, come evidenziato dall'aumento della perossidazione lipidica e dalle difese antiossidanti esaurite indotte dall'esposizione prolungata all'alluminio, sembra essere focalizzato nella corteccia prefrontale, nel cervelletto, nell'ippocampo e nel tronco cerebrale (Yuan et al., 2012 ; Kumar et al., 2011 ). È anche degno di nota il fatto che diversi autori abbiano riportato una relazione lineare tra l'aumento dei livelli cellulari dell'alluminio e le concentrazioni di proteine carbonili e proteine S100 (Mannello et al 2013 , Darbre et al 2013 , Darbre et al., 2011 ). Ciò è di particolare interesse in quanto queste molecole possono funzionare come DAMP e causare la stimolazione cronica dei PRR e quindi essere una fonte di attivazione immunitaria cronica come discusso sopra. Aumento dei livelli di perossidazione lipidica nel cervello con produzione di malondialdeide (MDA), 4-idrossi-2- transnononali (HNE o 4-idrossinonali (4-HNA)) e sostanze reattive agli acidi tiobarbituriche (TBARS) dopo somministrazione orale del cloruro di alluminio è anche una scoperta comune negli studi su piccoli animali (Newairy et al., 2009 ; Albendea et al., 2007 ; Yuan et al., 2012 ; Lu et al., 2013 ).
L'esposizione cronica dell'alluminio esercita anche profondi effetti nocivi sulle difese cellulari antiossidanti portando a livelli cellulari significativamente ridotti di glutatione transferasi, glutatione perossidasi, catalasi, superossido dismutasi e ridotto glutatione (GSH) (Nampoothiri et al., 2015 ; El-Demerdash 2004 ; Yousef 2004 ; Kumar et al., 2011 , Newairy et al., 2009 ). L'ingestione di alluminio riduce anche i livelli di GSH nei campioni di sangue umano (Khan et al., 2011 ). È interessante notare che l'alluminio diminuisce i livelli di questo tiolo inibendo l'attività dell'isocitrato deidrogenasi NADPH-dipendente nei mitocondri e l'enzima malico e l'isocitrato deidrogenasi NADPH nel citosol (Murakami e Yoshino 2004 ). Ciò è importante poiché i livelli impoveriti di questi enzimi rendono le cellule più sensibili alla perossidazione lipidica e al danno ossidativo del DNA mitocondriale dalle specie di ossigeno singoletto in un ambiente di stress ossidativo cronico (Kim e Park 2003 , Lee et al., 2002 ; Kochevar 2004 ). In questo contesto l'esistenza del danno ossidativo alle proteine mitocondriali e al DNA a seguito di un'esposizione prolungata all'alluminio è stata riportata da Sharma e colleghi (Sharma et al., 2013 ). L'ingestione di alluminio porta anche ad un aumento dello stress ossidativo, marcatori di perossidazione lipidica e diminuzione dei livelli di GSH nelle cellule epiteliali che rivestono l'intestino tenue (Orihuela et al., 2005 ). Questo esaurimento di GSH sembra essere influenzato dalla ridotta attività di GSH sintetasi, GSH reduttasi e cambiamenti non ancora eliminati alle membrane plasmatiche, con conseguente riduzione dell'afflusso di GSH dal lume alla mucosa (Orihuela et al., 2005 ). Questo esaurimento di GSH indotto dall'alluminio altera l'attività della calbindina-D9k con conseguente riduzione dell'assorbimento transcellulare degli ioni calcio (Orihuela et al., 2005 ). Dato il ruolo positivo giocato dagli ioni di calcio nel mantenimento dell'integrità della barriera epiteliale (Ma et al., Schepens et al., 2009 ), l'esaurimento di GSH potrebbe ben sostenere gli aumenti dell'infiammazione intestinale e della permeabilità della barriera intestinale causati dal prolungato consumo di alluminio (Pineton de Chambrun et al. 2014 ).
Esposizione dell'alluminio e sviluppo della disfunzione mitocondriale
Lo stress ossidativo e la conseguente disfunzione mitocondriale costituiscono il principale veicolo alla base della neurotossicità indotta da alluminio (per la revisione vedi Kumar e Gill 2014 ). L'esposizione agli ioni di alluminio porta ad una significativa diminuzione dell'attività del citocromo C ossidasi, NADH e succinato deidrogenasi e una successiva diminuzione dello stato 3 (stimolazione dell'ADP) e della respirazione mitocondriale dello stato 4, che sono probabilmente causate da cambiamenti conformazionali in questi enzimi come un risultato diretto del legame degli ioni di alluminio (Dua et al 2010 ; Mohan et al., 2009 ; Mustafa Rizvi et al., 2014 ; Kumar et al., 2008 ). L'alluminio altera anche la biogenesi mitocondriale diminuendo i livelli di attività del co-attivatore 1α (PGC-1α) del recettore attivato da proliferatore di perossisoma, direttamente o indirettamente come risultato dell'induzione di elevati livelli di stress ossidativo (Sharma et al., 2013 ). Gli ioni di alluminio mostrano anche la capacità di legarsi ai gruppi fosfato di ATP e ADP e inibiscono la fosforilazione di quest'ultima molecola e la defosforilazione del primo, e questo insieme alla capacità di inibire un'ampia gamma di enzimi chinasi e fosfatasi può gravemente compromettere omeostasi energetica (Kawahara e Kato-Negishi 2011 ).
L'alluminio può anche alterare la funzione mitocondriale indirettamente attraverso meccanismi come l'induzione dello stress del reticolo endoplasmatico (ER) (Mustafa Rizvi et al 2014 , Johnson et al., 2005 ). L'alluminio induce anche una disfunzione mitocondriale provocando il rilascio di ioni di calcio dalle riserve intracellulari, ed è interessante notare che il danno ossidativo indotto dall'alluminio e l'interruzione dell'omeostasi degli ioni calcio è simile nel modello a quello osservato nell'AD (Johnson et al., Walton 2012 ) . La relazione funzionale e fisica tra ER e mitocondri è ben documentata nel contesto dell'apoptosi, ma forse è poco discussa nel contesto dello stress ER che è sub-letale alla cellula (Vannuvel et al., 2013 ). In quest'ultimo ambiente, la risposta proteica dispiegata in generale, e l'attività della chinasi ER simile a RNA protein-chinasi in particolare, porta a uno stato di sottoperformance mitocondriale cronica piuttosto che a morte cellulare. Questa è un'area complessa e i lettori che desiderano approfondire tali meccanismi sono invitati a consultare il lavoro di Rainbolt et al. 2014 .
La dishomeostasi del calcio è ugualmente dannosa per la funzione mitocondriale e di vitale importanza nel mantenimento della funzione neurale abbinando la produzione di energia mitocondriale alla domanda (Rueda et al 2014 , Llorente-Folch et al., 2013 ). In particolare, aumenti modesti di ioni di calcio nel citosol in seguito all'aumento dell'attività nervosa agiscono come "pedale del gas" (o "acceleratore") per aumentare la produzionefattori di energia e mantenere l'omeostasi dell'ATP, quindi l'omeostasi del calcio compromessa può avere profondi effetti avversi sulla funzione neurale , anche in assenza di una sincera apoptosi (Gellerich et al., 2013 ) Gli effetti avversi dell'alluminio sull'omeostasi del calcio sono probabilmente un meccanismo coinvolto nella neuropatologia indotta dall'alluminio e ora consideriamo altri meccanismi per cui l'esposizione dell'alluminio potrebbe causare tipo di disfunzione astrocitica e microgliale osservata in molti bambini con una diagnosi confermata di ASD.
Gli effetti sopra riportati dell'alluminio sono riassunti in Fig. 1 .
Esposizione dell'alluminio e attivazione o disfunzione delle cellule gliali
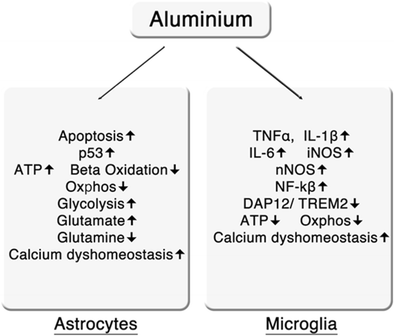
L'esposizione cronica o prolungata all'alluminio può indurre l'apoptosi degli astrociti con un meccanismo pensato per coinvolgere il DNA e il danno alla cromatina, e quindi mediato da p53 (Suarez-Fernandez et al., 1999 , Johnson et al., 2005 ). Un'altra via può comportare l'inibizione della funzione mitocondriale e la produzione di ATP, che alla fine causa necrosi, che può avere conseguenze neuroinfiammatorie profonde e prolungate (Lemire e Appanna 2011 ). L'esposizione prolungata all'alluminio può anche indurre cambiamenti metabolici significativi negli astrociti, che possono compromettere la funzionalità anche in assenza di degenerazione. Tali anomalie includono la riduzione dell'attività della γ-butirobetainealdeide deidrogenasi e della γ-butirrobetainealdeide diossigenasi e riduzione dei livelli di α-chetoglutarato (AKG), che porta a bassi livelli di L-carnitina e successivamente ridotta beta-ossidazione degli acidi grassi, disfunzione mitocondriale, ridotta produzione di ATP e aumento della lipogenesi (Mailloux et al., 2011 ; Lemire et al., 2011 ; Han et al., 2013 ).
L'aumentata lipogenesi successiva alla disfunzione mitocondriale indotta da alluminio attraverso questa via è consentita da significativi aumenti degli enzimi lipogenici come l'acetil CoA carbossilasi (Mailloux et al., 2006 ). L'aumento di attività di questi enzimi è accompagnato da una diminuzione dell'attività degli enzimi chiave all'interno della catena di trasporto degli elettroni e del ciclo dell'acido tricarbossilico (Kreb), come la succinato deidrogenasi e l'AKG, portando a una significativa diminuzione dei livelli di ATP prodotta da fosforilazione ossidativa (Mailloux et al., 2009 ; Mailloux et al., 2006 ; Mailloux et al., 2007 ). La perdita di attività di AKG nei mitocondri e nel citosol, probabilmente causata dal sequestro di questa molecola da parte delle difese antiossidanti, si traduce anche in conseguenze negative significative per la generazione di energia all'interno delle cellule gliali. In breve, AKG agisce per stabilizzare il fattore-1-alfa (HIF-1α) ipossia-inducibile nel citoplasma e ne previene la traslocazione nel nucleo. Tuttavia, in un ambiente cellulare con aumentati cationi di alluminio e successivamente ridotti livelli di AKG, HIF-1α si traslocava nel nucleo provocando aumenti nella trascrizione di esochinasi, piruvato chinasi, lattato deidrogenasi e glucosio-6-fosfato deidrogenasi, con l'effetto finale del passaggio dalla produzione di energia dalla fosforilazione ossidativa al percorso filogeneticamente più antico della produzione di energia attraverso la glicolisi (Mailloux e Appanna 2007 , Agrawal et al., 2007 ).
L'alluminio induce anche alterazioni significative del riciclo di glutammato / glutamina all'interno degli astrociti che porta ad un aumento della conversione di glutammina e glutammato, accoppiato con un aumento dell'assorbimento di glutammato e aumento dei livelli intracellulari di glutammina (Zielke et al., 1993 : Struys-Ponsar et al., 2000 ). Questo ha l'effetto di modulare la neurotrasmissione glutammatergica e GABA-ergica, ma può anche avere conseguenze bioenergetiche significative dato che l'aumento dei livelli di glutammato all'interno degli astrociti agisce come ulteriore stimolo per l'aumento della glicolisi (Albrecht et al., 2010 ; Bouzier-Sore e Pellerin 2013 ) . Queste osservazioni sono pertinenti dal punto di vista del neurosviluppo potenzialmente compromesso in quanto gli astrociti svolgono un ruolo importante nello sviluppo del cervello regolando i processi coinvolti nella trasmissione sinaptica, nella migrazione neuronale, nella sinaptogenesi e forse anche nella mielinizzazione (rivisto da Molofsky et al., 2012 ). Il peso dell'evidenza indica anche che l'attività di queste cellule gliali è di fondamentale importanza nello sviluppo e nel mantenimento di reti e circuiti neurali (Clarke and Barres 2013 ). Inoltre, vi sono ora prove considerevoli che indicano che la funzione alterata degli astrociti gioca un ruolo fondamentale nella patogenesi dei disturbi dello sviluppo neurologico (Molofsky et al., 2012 , Sloan and Barres 2013 , Yang et al., 2013 ).
L'alluminio può attivare la microglia che porta alla secrezione di TNF-α, IL-6 e ossido nitrico sintasi inducibile dalle citochine (iNOS o NOS-2) e lo sviluppo di PIC e ROS neuroinfiammatori (Bondy 2010 ; Zaky et al., 2013 ). Questa è anche una scoperta importante poiché ora esistono prove schiaccianti che dimostrano che la microglia svolge un ruolo indispensabile nello sviluppo del cervello regolando processi quali la potatura sinaptica, la plasticità sinaptica, la sinaptogenesi, lo sviluppo neuronale e altri processi vitali nella neurogenesi (Kettenmann et al. 2013 ). La disfunzione microgliale e / o l'innesco provocato da sfide immunitarie, eventi infiammatori o altri cambiamenti nel cervello che interferiscono con processi quali la potatura sinaptica e la proliferazione neurale sono ora considerati un importante ruolo causale nello sviluppo dell'ASD e di altri disturbi dello sviluppo neurologico come schizofrenia (Harry e Kraft 2012 ; Harry 2013 ; Hoeijmakers et al., 2016 ; Edmonson et al., 2016 ). Lo sviluppo graduale della microglia è regolato dall'attività di diversi geni e l'interruzione nell'espressione di questi geni può verificarsi a seguito di attivazione o disturbi immunitari prenatali nel microbiota (Matcovitch-Natan et al., 2016 ). L'alluminio può anche provocare attivazione e disfunzione microgliale attraverso diversi altri meccanismi, in particolare compromettendo la funzione del complesso molecolare multifunzionale della proteina adattatrice, proteina attivante il DNAX di 12 kD (DAP12) con TREM2, ovvero DAP12 / TREM2, che è espressa sul superficie delle cellule gliali (Bhattacharjee et al 2014 ; Zhao et al., 2014 ; Alexandrov et al., 2013 ).
Il complesso proteico DAP12 / TREM2 svolge un ruolo importante nella regolazione dell'omeostasi del sistema nervoso centrale (Paradowska-Gorycka e Jurkowska 2013 , Thrash et al., 2009 ). TREM2 lega una gamma di liposaccaridi, fosfolipidi e polianioni con la successiva attivazione di DAP12 e una serie di chinasi a valle che si accumulano nell'attivazione cellulare (Poliani et al., 2015 , Takahashi et al., 2007 ). L'attivazione dell'asse DAP12 / TREM2 ha un ruolo importante nel limitare i PIC e altre neurotossine in seguito all'attivazione dei TLR da parte di PAMP e DAMP e promuove la sopravvivenza e la proliferazione della microglia e di altre cellule della linea mieloide (Poliani et al., 2015 ; Peng et al 2013 ). Tuttavia, negli ultimi anni la maggior parte della ricerca sul complesso DAP12 / TREM2 si è concentrata sul suo ruolo nel consentire e regolare la fagocitosi microgliale (Neumann e Takahashi 2007 , Painter et al., 2015 ). TREM2 agisce come un sensore molecolare di detriti macromolecolari e svolge un ruolo indispensabile nella sua clearance immunogenicamente silente, risolvendo così le reazioni potenzialmente infiammatorie e agendo come un impedimento allo sviluppo della neuroinfiammazione (Painter et al., 2015 ; Takahashi et al., 2005 ; Takahashi et al., 2007 ; Hsieh et al., 2009 ).
I processi fagocitari della microglia svolgono un ruolo essenziale nella rimozione dei neuroni danneggiati o stressati e delle strutture sinaptiche, delle cellule apoptotiche e di altri detriti cellulari dal cervello, che hanno il potenziale di essere immunogenici e quindi di combattere lo sviluppo della neuroinfiammazione. Quindi, questo processo svolge un ruolo indispensabile nel mantenimento dell'omeostasi neurale (Fu et al., 2014 e Jones 2014 , Zhao e Lukiw 2013 ). Non sorprende che diversi gruppi di ricercatori abbiano riferito che il fallimento di questi incessanti processi fagocitari della microglia ha gravi conseguenze innate-immuni, pro-infiammatorie e neuropatogene (Jones et al., 2014 Koenigsknecht-Talboo et al., 2008 ). Fondamentalmente, come precedentemente notato, l'apoptosi microgliale è sia regolata che abilitata da TREM2 (Zhao e Lukiw 2013 ; Alexandrov et al., 2013 ; Jones et al., 2014 ; Wang et al., 2015 ). Compromissione nell'attività e / o espressione di TREM2 causa gravi menomazioni dell'attività fagocitaria della microglia che porta a un ampio spettro di patologie del sistema nervoso centrale tra cui neuroinfiammazione, perdita sinaptica, perdita neuronale e produzione esagerata di PIC (Wang et al., 2015 ; Hickman e El Khoury 2014 ; Jiang et al., 2014 ).
L'espressione di TREM2 è a sua volta regolata da un numero di miRNA, in particolare miR-34a (Zhao et al., 2013 ; Zhao e Lukiw 2013 ; Alexandrov et al., 2013 ). L'espressione microglial miR-34a è sovraregolata dall'attivazione del fattore di trascrizione pro-infiammatorio NF-κB (Zhang et al., 2011 ; Hickman and El Khoury 2014 ; Bhattacharjee et al., 2014 ). L'alluminio in forma adiuvante o ambientale sovra-regola NF-κB (Pogue et al., 2009 Bondy 2014 ), e quindi induce un numero di miRNA proinfiammatori sensibili alla NF-KB, in particolare miR-34a (Fu et al., 2014 , Jones et al 2014 ; Hickman e El Khoury 2014 ; Zhang et al., 2013 ; Zhao e Lukiw 2013 ), che a loro volta sottoregola l'espressione di TREM2 nelle membrane delle cellule gliali portando a un profondo deficit nella loro capacità fagocitica (Hickman and El Khoury 2014 ; Zhao et al., 2013 ; Bhattacharjee et al., 2014 ).
La sovraregolazione di NF-κB induce la sintesi di altri miRNA infiammatori come miR-9, miR-125b, miR-146a e miR-155, che sono riconosciuti driver di patologia in diverse malattie neurodegenerative come AD (Zhao et al., 2014 ; Lukiw 2012 ). Sebbene l'alluminio possa chiaramente provocare patologie croniche nelle persone con un'esposizione elevata o prolungata, è opportuno sottolineare ulteriormente che la probabilità che l'uso di adiuvanti possa essere la causa principale dell'ASD nei bambini, o addirittura dannosa per la grande maggioranza dei bambini con normali risposte immunitarie , sembra essere incredibilmente piccolo. Tuttavia, la situazione nei bambini con un sistema immunitario anormale e una predisposizione all'autoimmunità può essere diversa, e questa sarà l'ultima area di discussione in questo documento.
Le azioni dell'alluminio su astrociti e microglia sono riassunte in Fig. 2.
Sezione 4. Conseguenze dell'esposizione dell'alluminio in individui con risposte immunitarie anomale
Polimorfismi in geni immuno-effettore
Anomalie nelle prestazioni dei PPR sono sempre più associate allo sviluppo di condizioni neurodegenerative e autoimmuni (Moraes et al 2013 , Cook et al 2004 , Marshak-Rothstein 2006 , Pradhan et al 2012 , Liu et al 2014 ). In particolare, i polimorfismi funzionali nei geni che codificano le proteine coinvolte nell'effettuare la risposta immunitaria in seguito alla legatura di TLR-4, TLR-7 e TLR-9 aumentano la suscettibilità e / o la gravità di un range di neurologici, autoimmuni, infiammatori e malattie infettive, tra cui LES, artrite reumatoide, asma, sepsi ed epatite (Moraes et al., 2013 ; Dhaouadi et al., 2013 ; Fichna et al., 2016 ; Netea et al., 2012 ). Non sorprende che i polimorfismi funzionali nei geni delle citochine come quelli codificanti per TNF-α, IL-β, IL-6, IL-10 e IL-4 aumentano anche la suscettibilità a sviluppare una gamma di condizioni infiammatorie, infettive e autoimmuni (Morris e Berk 2015 ).
La presenza di questi polimorfismi può anche influenzare pesantemente la gravità e la traiettoria finale di queste malattie in un dato individuo (Arfanakis et al., 2013 : Tunçbilek 2014 ; Haukim et al., 2002 ; Hollegaard and Bidwell, 2006 ). Esistono anche evidenze che i polimorfismi funzionali dei geni che codificano le citochine e / oi loro recettori aumentano il rischio di patologie neuro-infiammatorie e neurodegenerative come ictus, AD e MS (Ottoboni et al., 2013 , Bagnoli et al., 2007 ; Hernandez e Baxter 2013 , Cui et al., 2012 ). I polimorfismi funzionali possono anche portare alla struttura alterata e alla funzione delle proteine recettore ed effettrici coinvolte in una vasta gamma di risposte inflammasome, e ancora una volta tali mutazioni sono anche coinvolte nella patogenesi di diverse malattie autoimmuni, neuro-immuni e neurodegenerative (Pontillo et al. 2012 ; Tan et al., 2013 , revisione di Yang e Chiang 2015 ). In generale, i polimorfismi "patologici" nei percorsi di localizzazione nucleare leucina-rich-repeat-protein (1 NLRP1) portano ad una risposta infiammatoria esagerata e / o prolungata (Levandowski et al., 2013 ).
È interessante notare che recenti ricerche hanno rivelato un'associazione significativa tra la presenza cumulativa di polimorfismi del gene immune e un aumentato rischio di sviluppare ASD (Ramos et al., 2012 ). Anche i polimorfismi a singolo nucleotide (SNP) nei geni immunitari in molti bambini mostrano uno schema caratteristico delle malattie autoimmuni come la SM (Jung et al., 2011 ). Non sorprende che i polimorfismi nel sistema HLA, in particolare HLA-DR4 e HLA-A2, siano associati a una probabilità significativamente maggiore di sviluppare la sindrome (Mostafa et al., 2013 , Torres et al., 2012 ). È anche degno di nota il fatto che la concomitante presenza di geni espressi in modo anomalo che regolano la regolazione e il rilascio di risposte immunitarie e infiammatorie può predire lo sviluppo di ASD con un'accuratezza dell'83% (Pramparo et al., 2015 ).
Vi sono ora abbondanti evidenze di espressione disregolata di geni che regolano la risposta immunitaria innata e adattativa nei bambini con ASD (Michel et al., 2012 ; Gupta et al., 2014 ). Esistono anche alcune prove di polimorfismi funzionali accoppiati con risposte PRR esagerate in almeno alcuni bambini affetti (Bennabi et al., 2015 , Enstrom et al., 2010 ). Esiste anche un corpo di ricerca che dimostra che le risposte immunitarie dopo l'attivazione dei TLR sono anormali in molti bambini che hanno ricevuto una diagnosi di ASD (Mead and Ashwood 2015 , Gesundheit et al 2013 , Enstrom et al., 2010 ). Inoltre, diversi gruppi di ricerca hanno fornito prove che indicano che la risposta immunitaria è anormale in molti bambini con ASD, e in particolare che i geni che regolano la regolazione e l'esecuzione delle risposte immunitarie e infiammatorie sono sovraregolati consentendo una risposta eccessiva e / o prolungata a un ambiente insulto (Voineagu e Eapen 2013 ; Koufaris e Sismani 2015 ). È anche degno di nota il fatto che diversi ricercatori abbiano anche segnalato un'associazione tra insulti infiammatori iniziali e il successivo sviluppo di disturbi immunitari cronici nei bambini con ASD (Siniscalco 2015 , Gottfried et al., 2015 , McDougle et al., 2015 ). Ciò è particolarmente degno di nota, poiché un'attivazione prolungata e / o eccessiva di percorsi immunitari e infiammatori che portano a stress ossidativo e nitrosativo può avere conseguenze dannose per l'integrità cellulare e tissutale, che a sua volta potrebbe portare all'attivazione cronica di percorsi immunitari e infiammatori e in definitiva di microglia e astrociti nel cervello come discuteremo ora.
Risposte immunitarie prolungate o eccessive e produzione di DAMP
Lo stress ossidativo attiva NFκB e altri fattori di trascrizione come la proteina-1 attivata (AP-1), portando alla sintesi e alla secrezione di PIC, varie specie di chemochine da parte di cellule di presentazione dell'antigene e attivazione e proliferazione di linfociti T e B (Morris et al. . 2015a ; Lucas et al., 2015 ). Tale attivazione e produzione di PIC porta alla sovraregolazione di iNOS e NADPH ossidasi, portando alla produzione di superossido, NO e perossinitrito, e quindi ulteriori aumenti nei livelli di ROS e di azoto reattivo (RNS) (Morris and Maes 2014 ; Morris et al. 2015a ). Questo rapporto bidirezionale e auto-amplificante tra lo sviluppo di un'infiammazione sistemica cronica, come evidenziato da un aumento dei livelli di PIC e dello stress nitrossidativo cronico come evidenziato da livelli aumentati di ROS e RNS, è spesso etichettato come un "ciclo autotossico" (Reuter et al 2010 ; Ortiz et al., 2013 ). Livelli eccessivi di ROS e RNS possono anche portare alla modificazione ossidativa e nitrosomatica di lipidi, proteine e DNA, portando a misfolding e conseguentemente immunogeniche, proteine, modificazione ossidativa del DNA e perossidazione lipidica delle membrane cellulari, insieme alla produzione di reattivi altamente reattivi chetoni e aldeidi (Lucas et al., 2015 ). Il risultato netto di questi processi è la formazione indiretta e diretta di DAMP in grado di attivare i recettori patogeni sulla superficie cellulare e nel citoplasma delle cellule immunitarie (Hwang 2013 ; Ortiz et al., 2013 ).
In sintesi, la presenza delle anomalie immunitarie discusse consente una risposta infiammatoria anormalmente eccessiva o prolungata in seguito all'esposizione ad adiuvanti di alluminio, portando alla produzione di DAMP e successiva attivazione immunitaria cronica in bambini geneticamente suscettibili. È interessante notare che diversi autori hanno segnalato la presenza di DAMP come proteina carbonile, MDA e la proteina box-1 (HMGB1) del gruppo di alta mobilità in almeno alcuni bambini hanno permesso una diagnosi di ASD, e tali DAMP potrebbero svolgere un ruolo importante in iniziare e mantenere uno stato di attivazione e infiammazione immunitaria osservato in molti bambini con una diagnosi di ASD (Babinska et al 2014 : Emanuele et al 2010 , Frank et al., 2016 , Napoli et al., 2013 ).
L'impegno cronico dei TLR da parte delle DAMP porta allo sviluppo di un ciclo di feedback positivo, aumentando il danno tissutale causato da PIC elevati, ROS e RNS e intensificando le risposte pro-infiammatorie, portando a uno stato di infiammazione cronica dello stress nitrossidativo, disfunzione mitocondriale attivazione di cellule gliali (Drexler e Foxwell 2010 , Piccinini e Midwood 2010 , Goh e Midwood 2012 , Morris e Berk 2015 ). Non sorprende che l'impegno cronico dei TLR, dei recettori simili ai recettori NIG (nucleeride binding colegion oligomerisation) e dei recettori RIG-simili sia implicato nella patogenesi e patofisiologia del LES, dell'artrite reumatoide e della SM (review (Drexler and Foxwell 2010 ; Piccinini and Midwood 2010 ; Goh e Midwood 2012 )). Pertinenza, la presenza di DAMP può anche portare all'attivazione cronica dell'infiammomaco (Anders and Schaefer 2014 ) che è anche implicato nello sviluppo di neuroinfiammazione e anormale segnalazione del sistema nervoso centrale caratteristico dei disordini neurodegenerativi e dello sviluppo neurologico (Tan et al. 2013 ; Singhal et al., 2014 ). E 'anche interessante che uno studio recente abbia riportato la presenza di complessi inflammasomi attivati in almeno alcuni bambini e ha fornito una diagnosi di ASD (Saresella et al., 2016 ). Vi è ora una prova schiacciante che l'attivazione prolungata e / o cronica di le vie infiammatorie provocano lo sviluppo di neuroinfiammazione cronica e / o priming microgliale e una breve spiegazione di questo processo costituirà la sottosezione finale di questo documento.
Microglia innescata attivazione immunitaria sistemica e neuro-infiammazione cronica
Esistono numerose prove che dimostrano che l'attivazione cronica del sistema immunitario e l'infiammazione sistemica possono portare allo sviluppo della neuro-infiammazione cronica (Perry and Holmes 2014 , Cunningham 2013 ). La comunicazione dei segnali infiammatori al cervello è mediata dai PIC attraverso una serie di vie, tra cui l'innervazione del nervo vago, il trasporto abilitato al trasporto attraverso la barriera emato-encefalica (BBB), l'attivazione delle cellule endoteliali all'interno del BBB e i macrofagi perivascolari, e infine attraverso il trasporto attraverso gli organi circumventricolari privi di un funzionale BBB (Morris and Maes 2012 , 2014 ). I segnali infiammatori trasdotti possono portare allo sviluppo della neuroinfiammazione cronica tramite l'attivazione della microglia se di intensità e / o durata sufficienti o portare allo sviluppo di microglia "innescata" (Perry and Holmes 2014 , Su e Federoff 2014 ). L'innesco microgliale comporta la sovraregolazione di una gamma di recettori di superficie come MHC classe II, CD11b e CD11c integrine, molecola co-stimolante CD86 e TLR-4 (Su e Federoff 2014 ).
Seguendo la sovraregolazione di questi recettori, tali microglia diventano squisitamente sensibili ai conseguenti stimoli infiammatori, portando ad una produzione esagerata di molecole neurotossiche che possono esacerbare la patologia preesistente e possono anche accelerare la progressione di malattie neuroinfiammatorie o neurodegenerative esistenti (Ferrari e Tarelli 2011 , Lunnon et al., 2011 ). Le microglia attivate esercitano i loro effetti neurotossici rilasciando i PIC, come TNF-α, IL-1β, IL-6 e IFN-γ, e i radicali liberi compreso il superossido, NO e perossinitrito, così come le molecole infiammatorie come la prostaglandina E2. Inoltre, TNF-α, IL-1β e IFN-γ possono agire come fonti secondarie di RNS e altre molecole infiammatorie agendo come potenti induttori di iNOS e attraverso la loro capacità di sovraregolare la cicloossigenasi-2 (COX-2) con la conseguente produzione di prostaglandina E2 (Morris et al. 2015b ; Perry et al., 2010 ). Il concetto di priming microgliale potrebbe cambiare il quadro di riferimento da una considerazione di una singola inoculazione contenente adiuvante di alluminio ad un effetto cumulativo causato da un programma di vaccinazione in cui successivi insulti immunitari in un breve periodo potrebbero provocare patologia cronica direttamente, provocando attività microgliale o, più indirettamente, provocando danni macromolecolari che potrebbero eventualmente raggiungere una soglia in grado di provocare patologia cronica. Va notato che vi è un accumulo di prove, anche se da studi su animali, che successivi e frequenti insulti immunitari e infiammatori postnatali giocano un ruolo fondamentale nell'avvento del priming microglial e nella genesi dei disordini dello sviluppo neurologico (Ibi e Yamada 2015 ; Nagai 2013 ). Esistono anche dati emergenti che implicano lo sviluppo del priming microgliale come fattore principale nello sviluppo di diverse se non tutte le malattie neurodegenerative (Bhattacharya et al., 2016 , Zhao et al., 2014 ; Shastri et al., 2013 ; Hoeijmakers et al., 2016 ) .
Sommario
La prova della neurotossicità dei cationi di alluminio (Al 3+ ) comprende: un'associazione tra esposizione cronica dell'alluminio e sviluppo di AD; il coinvolgimento di adiuvanti di alluminio nello sviluppo di ASIA; e prove epidemiologiche che indicano un'associazione tra l'uso di adiuvanti di alluminio e ASD. Esistono buone prove che suggeriscono che l'immunizzazione può accelerare o precipitare la transizione tra condizioni autoimmuni sintomatiche subcliniche e conclamate entro i primi 30 giorni post-immunizzazione, in particolare in quelli di età inferiore ai 50 anni. La risposta immunitaria alla immunizzazione può essere influenzata da variazioni nei geni HLA, TLR e citochine. Inoltre, l'esposizione dell'alluminio è associata alla produzione di citochine pro-infiammatorie e chemochine e allo sviluppo di stress ossidativo cronico, disfunzione mitocondriale e attivazione o disfunzione gliale; questi cambiamenti a loro volta sono associati all'ASD.
Conclusioni e direzioni future
L'alluminio non ha alcuna azione benefica fisiologica nota nel corpo umano e alcuni polimorfismi genetici predispongono a una maggiore suscettibilità ai suoi effetti avversi. Pertanto, si può fare un caso forte per evitare un'esposizione non necessaria a fonti ambientali di sali di alluminio, specialmente da parte di bambini, madri in gravidanza e donne in età fertile che possono iniziare una gravidanza. Tale evitamento non deve portare a disagi o disagi; le pentole in alluminio possono essere sostituite da alternative più sicure, mentre gli antitraspiranti contenenti alluminio, potenzialmente implicati nell'aumento dei casi di cancro al seno che interessano in particolare il quadrante superiore esterno della ghiandola mammaria, possono essere sostituiti da versioni non in alluminio. L'uso di sali di alluminio nei prodotti medici è un problema più controverso. Mentre gli antiacidi sono disponibili che non contengono sali di alluminio, la prevenzione di vaccinazioni che non contengono sali di alluminio come adiuvanti ha implicazioni politiche e finanziarie più ampie. Sembrerebbe prudente cercare di trovare un'alternativa ai coadiuvanti di alluminio il prima possibile ed eliminarne gradualmente l'uso.
Gli appunti
Ringraziamenti
Gli autori desiderano ringraziare Victoria Storey per il suo aiuto nella costruzione delle figure.
Conformità agli standard etici
Conflitto di interessi
Gli autori dichiarano di non avere conflitti di interesse.
Riferimenti
Abdallah MW, Hougaard DM, Norgaard-Pedersen B, Grove J, Bonefeld-Jorgensen EC, Mortensen EL (2012) Infezioni durante la gravidanza e dopo la nascita e il rischio di disturbi dello spettro autistico: uno studio basato sul registro che utilizza una coorte di nascita storica danese . Turk psikiyatri dergisi = Turkish J Psychiatry 23 (4): 229-235
Google Scholar
Abd-Elghaffar S, El-Sokkary GH, Sharkawy AA (2005) Neurotossicità indotta da alluminio e danno ossidativo nei conigli: effetto protettivo della melatonina. Neuro Endocrinol Lett 26 (5): 609-616
PubMed Google Scholar
Agmon-Levin N, Paz Z, israeliano E, Shoenfeld Y (2009) Vaccini e autoimmunità. Nat Rev Rheumatol 5 (11): 648-652. doi: 10.1038 / nrrheum.2009.196
PubMed CrossRef Google Scholar
Agmon-Levin N, Zafrir Y, Kivity S, Balofsky A, Amital H, Shoenfeld Y (2014) Sindrome da stanchezza cronica e fibromialgia in seguito all'immunizzazione con il vaccino contro l'epatite B: un altro angolo della sindrome autoimmunitaria (autoinfiammatoria) indotta da coadiuvanti '(ASIA). Immunol Res 60 (2-3): 376-383. doi: 10.1007 / s12026-014-8604-2
PubMed CrossRef Google Scholar
Agrawal A, Guttapalli A, Narayan S, Albert TJ, Shapiro IM, Risbud MV (2007) La stabilizzazione normossica di HIF-1alpha guida il metabolismo glicolitico e regola l'espressione genica di aggrecan in cellule polposo nucleo del disco intervertebrale di ratto. Am J Physiol Cell Physiol 293 (2): C621-C631. doi: 10.1152 / ajpcell.00538.2006
PubMed CrossRef Google Scholar
Ahmed SS, Santosh W (2010) Analisi della profilatura dei metalli e analisi della mappa di collegamento del morbo di Parkinson in fase iniziale: una nuova intuizione del marcatore dell'alluminio per la possibile diagnosi. PLoS One 5 (6): e11252. doi: 10.1371 / journal.pone.0011252
PubMed PubMedCentral CrossRef Google Scholar
Al-Ayadhi LY (2005) Metalli pesanti e oligoelementi in campioni di capelli di bambini autistici in Arabia Saudita centrale. Neurosci (Riyadh, Arabia Saudita) 10 (3): 213-218
Google Scholar
Albendea CD, Gomez-Trullen EM, Fuentes-Broto L, Miana-Mena FJ, Millan-Plano S, Reyes-Gonzales MC, Martinez-Ballarin E, Garcia JJ (2007) La melatonina riduce i danni ossidativi lipidici e proteici nei sinaptosomi dovuti all'alluminio . J Trace Elements Med Biol: Organo della società per minerali e oligoelementi (GMS) 21 (4): 261-268. doi: 10.1016 / j.jtemb.2007.04.002
CrossRef Google Scholar
Albrecht J, Sidoryk-Wegrzynowicz M, Zielinska M, Aschner M (2010) Ruoli di glutammina nella neurotrasmissione. Neuron Glia Biol 6 (4): 263-276. doi: 10.1017 / s1740925x11000093
PubMed CrossRef Google Scholar
Alexandrov PN, Zhao Y, Jones BM, Bhattacharjee S, Lukiw WJ (2013) L'espressione della proteina essenziale per la fagocitosi TREM2 è down-regolata da un miRNA-34a indotto da alluminio in una linea cellulare microgliale murina. J Inorg Biochem 128: 267-269. doi: 10.1016 / j.jinorgbio.2013.05.010
PubMed CrossRef Google Scholar
Al-Olayan EM, El-Khadragy MF, Abdel Moneim AE (2015) Le proprietà protettive della melatonina contro le lesioni neuronali indotte dall'alluminio. Int J Exp Pathol 96 (3): 196-202. doi: 10.1111 / iep.12122
PubMed PubMedCentral CrossRef Google Scholar
Anders HJ, Schaefer L (2014) Oltre i modelli molecolari associati al danno da lesioni tissutali, i recettori toll-like e gli inflammasomi guidano anche la rigenerazione e la fibrosi. J Am Soc Nephrol: JASN 25 (7): 1387-1400. doi: 10.1681 / asn.2014010117
PubMed PubMedCentral CrossRef Google Scholar
Andrews N, Miller E, Grant A, Stowe J, Osborne V, Taylor B (2004) Esposizione a Thimerosal in neonati e disturbi dello sviluppo: uno studio retrospettivo di coorte nel Regno Unito non supporta un'associazione causale. Pediatrics 114 (3): 584-591. doi: 10.1542 / peds.2003-1177-L
PubMed CrossRef Google Scholar
Arfanakis K, Fleischman DA, Grisot G, Barth CM, Varentsova A, Morris MC, Barnes LL, Bennett DA (2013) Infiammazione sistemica in anziani non demente Soggetti umani: Microstruttura e cognizione del cervello. PLoS One 8 (8): e73107. doi: 10.1371 / journal.pone.0073107
PubMed PubMedCentral CrossRef Google Scholar
Arnheim-Dahlstrom L, Pasternak B, Svanstrom H, Sparen P, Hviid A (2013) Eventi avversi tromboembolici autoimmuni, neurologici e venosi dopo l'immunizzazione di ragazze adolescenti con vaccino quadrivalente di papillomavirus umano in Danimarca e Svezia: studio di coorte. BMJ 347: f5906. doi: 10.1136 / bmj.f5906
PubMed PubMedCentral CrossRef Google Scholar
van Assen S, Elkayam O, Agmon-Levin N, Cervera R, Doran MF, Dougados M, Emery P, Geborek P, Ioannidis JP, Jayne DR, Kallenberg CG, Muller-Ladner U, Shoenfeld Y, Stojanovich L, Valesini G, Wulffraat NM, Bijl M (2011) Vaccinazione in pazienti adulti con malattie reumatiche infiammatorie autoimmuni: una revisione sistematica della letteratura per le raccomandazioni basate sull'evidenza della Lega europea contro l'reumatismo per la vaccinazione in pazienti adulti con malattie reumatiche infiammatorie autoimmuni. Autoimmun Rev 10 (6): 341-352. doi: 10.1016 / j.autrev.2010.12.003
PubMed CrossRef Google Scholar
Babinska K, Bucova M, Durmanova V, Lakatosova S, Janosikova D, Bakos J, Hlavata A, Ostatnikova D (2014) Aumento dei livelli plasmatici della proteina 1 scatola di gruppo ad alta mobilità (HMGB1) sono associati a un più alto punteggio di disfunzione gastrointestinale in individui con autismo. Physiol Res 63 (Suppl 4): S613-S618
PubMed Google Scholar
Bagavant H, Nandula SR, Kaplonek P, Rybakowska PD, Deshmukh US (2014) Alum, un adiuvante a base di alluminio, induce il disturbo di Sindrome di Sjögren nei topi. Clin Exp Rheumatol 32 (2): 251-255
PubMed PubMedCentral Google Scholar
Bagnoli S, Cellini E, Tedde A, Nacmias B, Piacentini S, Bessi V, Bracco L, Sorbi S (2007) Associazione del polimorfismo promotore IL10 nella malattia di Alzheimer in Italia. Neurosci Lett 418 (3): 262-265. doi: 10.1016 / j.neulet.2007.03.030
PubMed CrossRef Google Scholar
Basun H, Forssell LG, Wetterberg L, Winblad B (1991) Metalli e oligoelementi nel plasma e nel liquido cerebrospinale nel normale invecchiamento e nella malattia di Alzheimer. J Neural Transm Park Dis Dement Sect 3 (4): 231-258
PubMed Google Scholar
Beauchemin D, Kisilevsky R (1998) Un metodo basato su ICP-MS per l'analisi delle placche amiloidi di Alzheimer. Anal Chem 70 (5): 1026-1029
PubMed CrossRef Google Scholar
Bennabi M, Delorme R, Oliveira J, Fortier C, Lajnef M, Boukouaci W, Feugeas JP, Marzais F, Gaman A, Charron D, Ghaleh B, Krishnamoorthy R, Leboyer M, Tamouza R (2015) Dectin-1 Polymorphism: A Specificatore della malattia genetica nei disturbi dello spettro autistico? PLoS One 10 (9): e0137339. doi: 10.1371 / journal.pone.0137339
PubMed PubMedCentral CrossRef Google Scholar
Betancur C (2011) Eterogeneità etiologica nei disturbi dello spettro autistico: oltre 100 disordini genetici e genomici e ancora conteggio. Brain Res 1380: 42-77. doi: 10.1016 / j.brainres.2010.11.078
PubMed CrossRef Google Scholar
Bhattacharjee S, Zhao Y, Hill JM, Culicchia F, Kruck TPA, Percy ME, Pogue AI, Walton JR, Lukiw WJ (2013) Accumulo selettivo di alluminio nelle arterie cerebrali nella malattia di Alzheimer (AD). J Inorg Biochem 126: 35-37. doi: 10.1016 / j.jinorgbio.2013.05.007
PubMed PubMedCentral CrossRef Google Scholar
Deficit di Bhattacharjee S, Zhao Y, Lukiw WJ (2014) nel recettore-recettore per la fagocitosi del gene TREM2 endogeno miRNA-34a nella malattia di Alzheimer (AD); un aggiornamento. Aging frontale Neurosci 6: 116. doi: 10.3389 / fnagi.2014.00116
PubMed PubMedCentral Google Scholar
Bhattacharya A, Derecki NC, Lovenberg TW, Drevets WC (2016) Ruolo dei fattori neuro-immunologici nella patofisiologia dei disturbi dell'umore. Psychopharmacology 233 (9): 1623-1636. doi: 10.1007 / s00213-016-4214-0
PubMed CrossRef Google Scholar
Blaurock-Busch E, Amin OR, Rabah T (2011) Metalli pesanti e oligoelementi nei capelli e nelle urine di un campione di bambini arabi con disturbo dello spettro autistico. Maedica 6 (4): 247-257
PubMed PubMedCentral Google Scholar
Blaurock-Busch E, Amin OR, Dessoki HH, Rabah T (2012) Metodi tossici ed elementi essenziali nei capelli e gravità dei sintomi tra i bambini con autismo. Maedica 7 (1):
38-48 PubMed PubMedCentral Google Scholar
Bondy SC (2010) La neurotossicità dell'alluminio ambientale è ancora un problema. Neurotossicologia 31 (5): 575-581. doi: 10.1016 / j.neuro.2010.05.009
PubMed PubMedCentral CrossRef Google Scholar
Bondy SC (2014) L'esposizione prolungata a bassi livelli di alluminio porta a cambiamenti associati all'invecchiamento cerebrale e alla neurodegenerazione. Tossicologia 315: 1-7. doi: 10.1016 / j.tox.2013.10.008
PubMed CrossRef Google Scholar
Bouras C, Giannakopoulos P, Good PF, Hsu A, Hof PR, Perl DP (1997) A Laser Microprobe Analisi di massa del cervello Alluminio e ferro nella demenza pugilistica: confronto con la malattia di Alzheimer. Eur Neurol 38 (1): 53-58
PubMed CrossRef Google Scholar
Bouzier-Sore AK, Pellerin L (2013) Svelare la complessa natura metabolica degli astrociti. Front Cell Neurosci 7. doi: 10.3389 / fncel.2013.00179
Bushnell PJ (2013) Numero speciale: influenze ambientali e meccanismi emergenti nell'eziologia dell'autismo. Neurotoxicol Teratol 36: 1-2. doi: 10.1016 / j.ntt.2013.03.001
PubMed CrossRef Google Scholar
Careaga M, Ashwood P (2012) Disturbi dello spettro autistico: dall'immunità al comportamento. Metodi Mol Biol 934: 219-240. doi: 10.1007 / 978-1-62703-071-7_12
PubMed CrossRef Google Scholar
Castiblanco J, Anaya JM (2015) Genetica e vaccini nell'era della medicina personalizzata. Current Genomics 16 (1): 47-59. doi: 10.2174 / 1389202916666141223220551
PubMed PubMedCentral CrossRef Google Scholar
Cerpa-Cruz S, Paredes-Casillas P, Landeros Navarro E, Bernard-Medina AG, Martinez-Bonilla G, Gutierrez-Urena S (2013) Eventi avversi dopo l'immunizzazione con vaccini contenenti adiuvanti. Immunol Res 56 (2-3): 299-303. doi: 10.1007 / s12026-013-8400-4
PubMed CrossRef Google Scholar
Chafi AH, Hauw JJ, Rancurel G, Berry JP, Galle C (1991) Assenza di alluminio nel tessuto cerebrale del morbo di Alzheimer: studi di microscopia elettronica e microscopia ionica. Neurosci Lett 123 (1): 61-64
PubMed CrossRef Google Scholar
Chao C, Klein NP, Velicer CM, Sy LS, Slezak JM, Takhar H, Ackerson B, Cheetham TC, Hansen J, Deosaransingh K, Emery M, Liaw KL, Jacobsen SJ (2012) Sorveglianza delle condizioni autoimmuni in seguito all'uso di routine del quadrivalente vaccino contro il papillomavirus umano. J Intern Med 271 (2): 193-203. doi: 10.1111 / j.1365-2796.2011.02467.x
PubMed CrossRef Google Scholar
Chen S, Li Z, He Y, Zhang F, Li H, Liao Y, Wei Z, Wan G, Xiang X, Hu M, Xia K, Chen X, Tang J (2015) Elevato numero di copie del DNA mitocondriale nelle cellule del sangue periferico è associato all'autismo infantile. BMC Psychi 15:50. doi: 10.1186 / s12888-015-0432-y
CrossRef Google Scholar
Clarke LE, Barres BA (2013) Ruoli emergenti degli astrociti nello sviluppo del circuito neurale. Nat Rev Neurosci 14 (5): 311-321. doi: 10.1038 / nrn3484
PubMed PubMedCentral CrossRef Google Scholar
Cook DN, Pisetsky DS, Schwartz DA (2004) Recettori toll-like nella patogenesi della malattia umana. Nat Immunol 5 (10): 975-979
PubMed CrossRef Google Scholar
Cruz-Tapias P, Agmon-Levin N, israeliana E, Anaya JM, Shoenfeld Y (2013) Sindrome autoimmune (autoinfiammatoria) indotta da adiuvanti (ASIA) - modelli animali come prova del concetto. Current Med Chem 20 (32): 4030-4036
CrossRef Google Scholar
Cui G, Wang H, Li R, Zhang L, Li Z, Wang Y, Hui R, Ding H, Wang DW (2012) Polimorfismo del promotore del gene alfa (TNF-alfa) del fattore di necrosi tumorale, livello di TNF-alfa circolante, e fattore di rischio cardiovascolare per ictus ischemico. J Neuroinfiammazione 9: 235. doi: 10.1186 / 1742-2094-9-235
PubMed PubMedCentral CrossRef Google Scholar
Cunningham C (2013) Microglia e neurodegenerazione: il ruolo dell'infiammazione sistemica. Glia 61: 71-90
PubMed CrossRef Google Scholar
Darbre PD, Pugazhendhi D, Mannello F (2011) Malattie dell'alluminio e del seno umano. J Inorg Biochem 105 (11): 1484-1488. doi: 10.1016 / j.jinorgbio.2011.07.017
PubMed CrossRef Google Scholar
Darbre PD, Mannello F, Exley C (2013) Cancro dell'alluminio e della mammella: Fonti di esposizione, misure tissutali e meccanismi di azioni tossicologiche sulla biologia mammaria. J Inorg Biochem 128: 257-261. doi: 10.1016 / j.jinorgbio.2013.07.005
PubMed CrossRef Google Scholar
De Sole P, Rossi C, Chiarpotto M, Ciasca G, Bocca B, Alimonti A, Bizzarro A, Rossi C, Masullo C (2013) Possibile relazione tra complesso Al / ferritina e morbo di Alzheimer. Clin Biochem 46 (1-2): 89-93. doi: 10.1016 / j.clinbiochem.2012.10.023
PubMed CrossRef Google Scholar
Depino AM (2013) Infiammazione periferica e centrale nei disturbi dello spettro autistico. Mol Cell Neurosci 53: 69-76. doi: 10.1016 / j.mcn.2012.10.003
PubMed CrossRef Google Scholar
DeStefano F (2007) Vaccini e autismo: le prove non supportano un'associazione causale. Clin Pharmacol Ther 82 (6): 756-759. doi: 10.1038 / sj.clpt.6100407
PubMed CrossRef Google Scholar
Dhaouadi T, Sfar I, Haouami Y, Abdelmoula L, Turki S, Hassine L (2013) Polimorfismi del recettore Toll-4 e CD14 nel lupus eritematoso sistemico e nell'artrite reumatoide. Biomark Res 1:20
PubMed PubMedCentral CrossRef Google Scholar
Drexler SK, Foxwell BM (2010) Il ruolo dei recettori toll-like nell'infiammazione cronica. Int J Biochem Cell Biol 42 (4): 506-518. doi: 10.1016 / j.biocel.2009.10.009
PubMed CrossRef Google Scholar
Dua R, Sunkaria A, Kumar V, Gill KD (2010) Metabolismo energetico mitocondriale alterato e proprietà cinetiche della citocromo ossidasi a seguito dell'esposizione acuta al fosfuro di alluminio nel fegato di ratto. Food Chem Toxicol 48 (1): 53-60. doi: 10.1016 / j.fct.2009.09.014
PubMed CrossRef Google Scholar
Edmonson CA, Ziats MN, Rennert OM (2016) Un ruolo non infiammatorio per la microglia nei disturbi dello spettro autistico. Front Neurol 7: 9. doi: 10.3389 / fneur.2016.00009
PubMed PubMedCentral CrossRef Google Scholar
Edwardson JA, Klinowski J, Oakley AE, Perry RH, Candy JM (1986) Aluminosilicati e cervello che invecchia: implicazioni per la patogenesi della malattia di Alzheimer. CIBA Found Symp 121: 160-179
PubMed Google Scholar
Eisenbarth SC, Colegio OR, O'Connor W, Sutterwala FS, Flavell RA (2008) Ruolo cruciale per l'inflammasoma Nalp3 nelle proprietà immunostimolatorie degli adiuvanti di alluminio. Nature 453 (7198): 1122-1126. doi: 10.1038 / nature06939
PubMed PubMedCentral CrossRef Google Scholar
Eisenkraft A, Falk A, Finkelstein A (2013) Il ruolo del glutammato e del sistema immunitario nel danno al SNC indotto dall'organofosfato. Neurotox Res 24 (2): 265-279. doi: 10.1007 / s12640-013-9388-1
PubMed CrossRef Google Scholar
El-Demerdash FM (2004) Effetto antiossidante di vitamina E e selenio sulla perossidazione lipidica, attività enzimatica e parametri biochimici nei ratti esposti all'alluminio. J Trace Elem Med Biol 18 (1): 113-121. doi: 10.1016 / j.jtemb.2004.04.001
PubMed CrossRef Google Scholar
Emanuele E, Boso M, Brondino N, Pietra S, Barale F, Ucelli di Nemi S, Politi P (2010) Aumento dei livelli sierici di proteine del gruppo 1 ad alta mobilità in pazienti con disturbo autistico. Prog Neuro-Psychopharmacol Biol Psychiatry 34 (4): 681-683. doi: 10.1016 / j.pnpbp.2010.03.020
CrossRef Google Scholar
Enstrom AM, Onore CE, Van de Water JA, Ashwood P (2010) Le risposte dei monociti differenziali ai ligandi TLR nei bambini con disturbi dello spettro autistico. Brain Behav Immun 24 (1): 64-71. doi: 10.1016 / j.bbi.2009.08.001
PubMed CrossRef Google Scholar
Esposito S, Prada E, Mastrolia MV, Tarantino G, Codeca C, Rigante D (2014) Sindrome autoimmune / infiammatoria indotta da adiuvanti (ASIA): indizi e insidie nel contesto pediatrico. Immunol Res 60 (2-3): 366-375. doi: 10.1007 / s12026-014-8586-0
PubMed CrossRef Google Scholar
Estes ML, McAllister AK (2015) Mediatori immunitari nel cervello e nei tessuti periferici nel disturbo dello spettro autistico. Nat Rev Neurosci 16 (8): 469-486. doi: 10.1038 / nrn3978
PubMed CrossRef Google Scholar
Exley C (2005) L'ipotesi della cascata di alluminio-amiloide e il morbo di Alzheimer. Subcell Biochem 38: 225-234
PubMed CrossRef Google Scholar
Exley C, Esiri MM (2006) Grave angiopatia cerebrale congestizia coincidente con un aumento dell'alluminio del cervello in un residente di Camelford, Cornovaglia, Regno Unito. J Neurol Neurosurg Psychiatry 77 (7): 877-879. doi: 10.1136 / jnnp.2005.086553
PubMed PubMedCentral CrossRef Google Scholar
Exley C, Vickers T (2014) Elevato contenuto di alluminio cerebrale e malattia di Alzheimer ad esordio precoce in un individuo esposto professionalmente all'alluminio: un caso clinico. J Med Case Rep 8 (1): 41
PubMed PubMedCentral CrossRef Google Scholar
Exley C, Mamutse G, Korchazhkina O, Pye E, Strekopytov S, Polwart A, Hawkins C (2006) Elevata escrezione urinaria di alluminio e ferro nella sclerosi multipla. Mult Scler 12 (5): 533-540
PubMed CrossRef Google Scholar
Ferrari C, Tarelli R (2011) Malattia di Parkinson e infiammazione sistemica. Parkinsons Dis 2011: 436813
PubMed PubMedCentral Google Scholar
Fichna M, Zurawek M, Fichna P, Januszkiewicz-Lewandowska D, Ruchala M, Nowak J (2016) Polimorfismi del gene Toll-Like Receptor-3 in insufficienza surrenalica autoimmune e diabete di tipo 1 in pazienti polacchi. Arch Immunol Ther Exp 64 (1): 83-87. doi: 10.1007 / s00005-015-0360-z
CrossRef Google Scholar
Fido A, Al-Saad S (2005) Tracce tossiche nei capelli dei bambini con autismo. Autismo 9 (3): 290-298. doi: 10.1177 / 1362361305053255
PubMed CrossRef Google Scholar
Flashner BM, Russo ME, Boileau JE, Leong DW, Gallicano GI (2013) Fattori epigenetici e disturbi dello spettro autistico. NeuroMolecular Med 15 (2): 339-350. doi: 10.1007 / s12017-013-8222-5
PubMed CrossRef Google Scholar
Flaten TP (2001) L'alluminio come fattore di rischio nella malattia di Alzheimer, con particolare attenzione all'acqua potabile. Brain Res Bull 55 (2): 187-196
PubMed CrossRef Google Scholar
Franchi L, Nunez G (2008) L'inflammasoma Nlrp3 è fondamentale per la secrezione di IL-1beta mediata dall'idrossido di alluminio, ma non è adatta per l'attività adiuvante. Eur J Immunol 38 (8): 2085-2089. doi: 10.1002 / eji.200838549
PubMed PubMedCentral CrossRef Google Scholar
Frank MG, Weber MD, Fonken LK, Hershman SA, Watkins LR, Maier SF (2016) Lo stato redox dell'allarme in HMGB1 è un fattore chiave nell'innesco neuroinfiammatorio e microgliale: un ruolo per l'inflammasoma NLRP3. Brain Behav Immun 55: 215-224. doi: 10.1016 / j.bbi.2015.10.009
PubMed CrossRef Google Scholar
Frustaci A, Neri M, Cesario A, Adams J, Domenici E, Dalla Bernardina B (2012) Biomarcatori correlati allo stress ossidativo nell'autismo: revisione sistematica e meta-analisi. Gratuito Radic Biol Med 52: 2128-2141
PubMed CrossRef Google Scholar
Fu R, Shen Q, Xu P, Luo JJ, Tang Y (2014) Fagocitosi della microglia nelle malattie del sistema nervoso centrale. Mol Neurobiol 49 (3): 1422-1434. doi: 10.1007 / s12035-013-8620-6
PubMed PubMedCentral CrossRef Google Scholar
Fulgenzi A, Vietti D, Ferrero ME (2014) Coinvolgimento dell'alluminio in neurotossicità. Biomed Res Int 2014: 758323. doi: 10.1155 / 2014/758323
PubMed PubMedCentral Google Scholar
Gellerich FN, Gizatullina Z, Gainutdinov T, Muth K, Seppet E, Orynbayeva Z, Vielhaber S (2013) Il controllo dell'energetica mitocondriale del cervello da parte del calcio citosolico: il pedale del gas mitocondriale. IUBMB Life 65 (3): 180-190. doi: 10.1002 / iub.1131
PubMed CrossRef Google Scholar
Georgiades S, Szatmari P, Boyle M (2013) Importanza dello studio dell'eterogeneità nell'autismo. Neuropsychiatry 3 (2): 123-125. doi: 10.2217 / npy.13.8
CrossRef Google Scholar
Gesundheit B, Rosenzweig JP, Naor D, Lerer B, Zachor DA, Prochazka V, Melamed M, Kristt DA, Steinberg A, Shulman C, Hwang P, Koren G, Walfisch A, Passweg JR, Snowden JA, Tamouza R, Leboyer M , Farge-Bancel D, Ashwood P (2013) Considerazioni immunologiche e autoimmuni dei disturbi dello spettro autistico. J Autoimmun 44: 1-7. doi: 10.1016 / j.jaut.2013.05.005
PubMed CrossRef Google Scholar
Goh FG, Midwood KS (2012) Pericolo intrinseco: attivazione dei recettori Toll-like nell'artrite reumatoide. Reumatologia (Oxford) 51 (1): 7-23. doi: 10.1093 / reumatologia / ker257
CrossRef Google Scholar
Goh S, Dong Z, Zhang Y, DiMauro S, Peterson BS (2014) Disfunzione mitocondriale come sottotipo neurobiologico del disturbo dello spettro autistico: evidenze dall'imaging cerebrale. JAMA Psychiatry 71 (6): 665-671. doi: 10.1001 / jamapsychiatry.2014.179
PubMed PubMedCentral CrossRef Google Scholar
Good PF, Perl DP, Bierer LM, Schmeidler J (1992) Accumulo selettivo di alluminio e ferro nei grovigli neurofibrillari del morbo di Alzheimer: uno studio di microscopia laser (LAMMA). Ann Neurol 31 (3): 286-292
PubMed CrossRef Google Scholar
Gottfried C, Bambini-Junior V, Francis F, Riesgo R, Savino W (2015) L'impatto delle alterazioni neuroimmune nel disturbo dello spettro autistico. Front Psychiatry 6: 121. doi: 10.3389 / fpsyt.2015.00121
PubMed PubMedCentral CrossRef Google Scholar
Grether JK, Rosen NJ, Smith KS, Croen LA (2009) Indagine sui cambiamenti nella segnalazione di autismo nel Dipartimento per i Servizi di Sviluppo della California. J Autism Dev Disord 39 (10): 1412-1419. doi: 10.1007 / s10803-009-0754-z
PubMed CrossRef Google Scholar
Grimaldi-Bensouda L, Guillemot D, Godeau B, Benichou J, Lebrun-Frenay C, Papeix C, Labauge P, Berquin P, Penfornis A, Benhamou PY, Nicolino M, Simon A, Viallard JF, Costedoat-Chalumeau N, Courcoux MF , Pondarre C, Hilliquin P, Chatelus E, Foltz V, Guillaume S, Rossignol M, Abenhaim L (2014) Disordini autoimmuni e vaccinazione quadrivalente di papillomavirus umano di soggetti giovani donne. J Intern Med 275 (4): 398-408. doi: 10.1111 / joim.12155
PubMed CrossRef Google Scholar
Guevara-Campos J, Gonzalez-Guevara L, Puig-Alcaraz C, Cauli O (2013) Disturbi dello spettro autistico associati a una carenza degli enzimi della catena respiratoria mitocondriale. Metab Brain Dis 28 (4): 605-612. doi: 10.1007 / s11011-013-9419-x
PubMed CrossRef Google Scholar
Guimaraes LE, Baker B, Perricone C, Shoenfeld Y (2015) Vaccini, adiuvanti e autoimmunità. Pharmacol Res 100: 190-209. doi: 10.1016 / jrsrs.2015.08.003
PubMed CrossRef Google Scholar
Gupta S, Ellis SE, Ashar FN, Moes A, Bader JS, Zhan J, West AB, Arking DE (2014) L'analisi del trascrittoma rivela la disregolazione dei geni di risposta immunitaria innata e dei geni dipendenti dall'attività neuronale nell'autismo. Nat Commun 5. doi: 10.1038 / ncomms6748
Hadjkacem I, Ayadi H, Turki M, Yaich S, Khemekhem K, Walha A, Cherif L, Moalla Y, Ghribi F (2016) fattori prenatali, perinatali e postnatali associati al disturbo dello spettro autistico. J Pediatr 92 (6): 595-601. doi: 10.1016 / j.jped.2016.01.012
CrossRef Google Scholar
Hallmayer J, Cleveland S, Torres A et al (2011) ereditabilità genetica e fattori ambientali condivisi tra coppie gemellate con autismo. Arch Gen Psychiatry 68 (11): 1095-1102. doi: 10.1001 / archgenpsychiatry.2011.76
PubMed PubMedCentral CrossRef Google Scholar
Han S, Lemire J, Appanna VP, Auger C, Castonguay Z, Appanna VD (2013) Come alluminio, un generatore di ROS intracellulare promuove malattie epatiche e neurologiche: il racconto metabolico. Cell Biol Toxicol 29 (2): 75-84. doi: 10.1007 / s10565-013-9239-0
PubMed CrossRef Google Scholar
Haralambieva IH, Ovsyannikova IG, Pankratz VS, Kennedy RB, Jacobson RM, Polonia GA (2013) La base genetica per la variazione interindividuale della risposta immunitaria al vaccino contro il morbillo: nuova comprensione e nuovi approcci vaccinali. Exp Rev Vacc 12 (1): 57-70. doi: 10.1586 / erv.12.134
CrossRef Google Scholar
Harry GJ (2013) Microglia durante lo sviluppo e l'invecchiamento. Pharmacol Ther 139 (3): 313-326. doi: 10.1016 / j.pharmthera.2013.04.013
PubMed PubMedCentral CrossRef Google Scholar
Harry GJ, Kraft AD (2012) Microglia nel cervello in via di sviluppo: un potenziale bersaglio con effetti a vita. Neurotoxicology 33 (2): 191-206. doi: 10.1016 / j.neuro.2012.01.012
PubMed PubMedCentral CrossRef Google Scholar
Haukim N, Bidwell JL, Smith AJP, Keen LJ, Gallagher G, Kimberly R, Huizinga T, McDermott MF, Oksenberg J, McNicholl J, Pociot F, Hardt C, Dalfonso S (2002) Polimorfismo del gene delle citochine nella malattia umana: on- line databases, Supplement 2. Genes Immun 3 (6): 313-330
PubMed CrossRef Google Scholar
Herbert MR (2010) Contributi dell'ambiente e della fisiologia ambientalmente vulnerabile ai disturbi dello spettro autistico. Curr Opin Neurol 23 (2): 103-110. doi: 10.1097 / WCO.0b013e328336a01f
PubMed CrossRef Google Scholar
Hertz-Picciotto I (2009) Commento: cambiamento diagnostico e aumento della prevalenza dell'autismo. Int J Epidemiol 38 (5): 1239-1241. doi: 10.1093 / ije / dyp258
PubMed PubMedCentral CrossRef Google Scholar
Hickman SE, El Khoury J (2014) TREM2 e la neuroimmunologia del morbo di Alzheimer. Biochem Pharmacol 88 (4): 495-498. doi: 10.1016 / j.bcp.2013.11.021
PubMed CrossRef Google Scholar
Hoeijmakers L, Heinen Y, Van Dam AM, Lucassen PJ, Korosi A (2016) Microglial Priming e Alzheimer: un possibile ruolo per le (prime) sfide immunitarie e l'epigenetica? Front Hum Neurosci 10: 398. doi: 10.3389 / fnhum.2016.00398
PubMed PubMedCentral CrossRef Google Scholar
Hollegaard MV, Bidwell JL (2006) Polimorfismo del gene delle citochine nelle malattie umane: database on-line, supplemento 3. Genes Immun 7 (4): 269-276
PubMed CrossRef Google Scholar
Hooker BS (2014) Tempi di vaccinazione contro il parotite e la rosolia e autismo tra i giovani ragazzi afro-americani: una rianalisi dei dati CDC. Transl Neurodegener 3 (1): 1-6. doi: 10.1186 / 2047-9158-3-16
CrossRef Google Scholar
Hornung V, Bauernfeind F, Halle A, Samstad EO, Kono H, Rock KL, Fitzgerald KA, Latz E (2008) I cristalli di silice e i sali di alluminio attivano l'inflammasoma NALP3 attraverso la destabilizzazione fagosomiale. Nat Immunol 9 (8): 847-856. doi: 10.1038 / ni.1631
PubMed PubMedCentral CrossRef Google Scholar
Hsieh CL, Koike M, Spusta SC, Niemi EC, Yenari M, Nakamura MC, Seaman WE (2009) Un ruolo per i ligandi TREM2 nella fagocitosi delle cellule neuronali apoptotiche mediante microglia. J Neurochem 109 (4): 1144-1156. doi: 10.1111 / j.1471-4159.2009.06042.x
PubMed PubMedCentral CrossRef Google Scholar
Hwang O (2013) Ruolo dello stress ossidativo nella malattia di Parkinson. Exp Neurobiol 22: 11-17
PubMed PubMedCentral CrossRef Google Scholar
Ibi D, Yamada K (2015) Obiettivi terapeutici per i disturbi del neurosviluppo emergenti da modelli animali con attivazione immunitaria perinatale. Int J Mol Sci 16 (12): 28218-28229. doi: 10.3390 / ijms161226092
PubMed PubMedCentral CrossRef Google Scholar
Israeliana (2012) sindrome da guerra del Golfo come parte della sindrome autoimmune (autoinfiammatoria) indotta da adiuvante (ASIA). Lupus 21 (2): 190-194. doi: 10.1177 / 0961203311429552
PubMed CrossRef Google Scholar
Jacobs RW, Duong T, Jones RE, Trapp GA, Scheibel AB (1989) Un riesame dell'alluminio nella malattia di Alzheimer: analisi mediante microscopia a dispersione di energia a raggi X e spettrofotometria di assorbimento atomico senza fiamma. Can J Neurol Sci 16: 498-503
PubMed CrossRef Google Scholar
Jensen-Jarolim E (2015) Allergy in Allergy and Allergen immunotherapy. World Aller Org J 8 (1): 7. doi: 10.1186 / s40413-015-0060-5
Google Scholar
Jiang T, Tan L, Zhu XC, Zhang QQ, Cao L, Tan MS, Gu LZ, Wang HF, Ding ZZ, Zhang YD, Yu JT (2014) Upregulation di TREM2 migliora la neuropatologia e salva deficit cognitivi spaziali in un modello di topo transgenico della malattia di Alzheimer. Neuropsychopharmacology 39 (13): 2949-2962. doi: 10.1038 / npp.2014.164
PubMed PubMedCentral CrossRef Google Scholar
Johnson VJ, Kim SH, Sharma RP (2005) alluminio-maltolato induce apoptosi e necrosi nelle cellule neuro-2a: ruolo potenziale per la segnalazione p53. Toxicol Sci 83 (2): 329-339. doi: 10.1093 / toxsci / kfi028
PubMed CrossRef Google Scholar
Jones BM, Bhattacharjee S, Dua P, Hill JM, Zhao Y, Lukiw WJ (2014) Regolazione dell'amiloidogenesi attraverso il recettore di attivazione naturale espresso in cellule mieloidi / microgliali 2 (TREM2). Front Cell Neurosci 8:94. doi: 10.3389 / fncel.2014.00094
PubMed PubMedCentral Google Scholar
Jung JY, Kohane IS, Wall DP (2011) Identificazione delle firme geniche autoimmuni nell'autismo. Transl Psychiatry 1: e63. doi: 10.1038 / tp.2011.62
PubMed PubMedCentral CrossRef Google Scholar
Kawahara M (2005) Effetti dell'alluminio sul sistema nervoso e il suo possibile legame con le malattie neurodegenerative. J Alzheimer's Dis: JAD 8 (2): 171-182 discussione 209-115
PubMed CrossRef Google Scholar
Kawahara M, Kato-Negishi M (2011) Legame tra l'alluminio e la patogenesi della malattia di Alzheimer: l'integrazione delle ipotesi di cascata di alluminio e amiloide. Int J Alzheimers Dis 2011: 276393. doi: 10.4061 / 2011/276393
PubMed PubMedCentral Google Scholar
Kern JK, Geier DA, Sykes LK, Haley BE, Geier MR (2016) La relazione tra mercurio e autismo: una revisione e una discussione completa. J Trace Elem Med Biol 37: 8-24. doi: 10.1016 / j.jtemb.2016.06.002
PubMed CrossRef Google Scholar
Kettenmann H, Kirchhoff F, Verkhratsky A (2013) Microglia: nuovi ruoli per lo spogliarellista sinaptico. Neurone 77 (1): 10-18. doi: 10.1016 / j.neuron.2012.12.023
PubMed CrossRef Google Scholar
Khan H, Khan MF, Jan SU, Ullah N (2011) Effetto del metallo di alluminio sul livello di glutatione (GSH) nel plasma e nella frazione citosolica del sangue umano. Pak J Pharm Sci 24 (1): 13-18
PubMed Google Scholar
Kim SY, Park JW (2003) Difesa cellulare contro il danno ossidativo indotto dall'ossigeno singoletto con NADP + isocitrato deidrogenasi citosolico. Free Radic Res 37 (3): 309-316
PubMed CrossRef Google Scholar
Kim YS, Leventhal BL, Koh YJ, Fombonne E, Laska E, Lim CE, Cheon KA, Kim SJ, Kim YK, Lee H, Song DH, Grinker RR (2011) Prevalenza dei disturbi dello spettro autistico in un campione di popolazione totale. Am J Psychiatry 168 (9): 904-912. doi: 10.1176 / appi.ajp.2011.10101532
PubMed CrossRef Google Scholar
King M, Bearman P (2009) Cambiamenti diagnostici e aumento della prevalenza di autismo. Int J Epidemiol 38 (5): 1224-1234. doi: 10.1093 / ije / dyp261
PubMed PubMedCentral CrossRef Google Scholar
Klei L, Sanders SJ, Murtha MT, Hus V, Lowe JK, Willsey AJ, Moreno-De-Luca D, Yu TW, Fombonne E, Geschwind D, Grice DE, Ledbetter DH, Lord C, Mane SM, Martin CL, Martin DM, Morrow EM, Walsh CA, Melhem NM, Chaste P, Sutcliffe JS, Stato MW, Cook EH Jr, Roeder K, Devlin B (2012) Le varianti genetiche comuni, che agiscono additivamente, sono una delle principali fonti di rischio per l'autismo. Autismo molecolare 3 (1): 9. doi: 10.1186 / 2040-2392-3-9
PubMed PubMedCentral CrossRef Google Scholar
Klein C, Lohmann K (2009) Malattia di Parkinson: la "malattia di Parkinson" è un'entità clinica distinta? Neurologia 72 (2): 106-107. doi: 10.1212 / 01.wnl.0000333666.65522.8d
PubMed CrossRef Google Scholar
Kochevar IE (2004) Segnalazione dell'ossigeno singoletto: dall'intimo al globale. Sci STKE 2004 (221): pe7. doi: 10.1126 / stke.2212004pe7
Koenigsknecht-Talboo J, Meyer-Luehmann M, Parsadanian M, Garcia-Alloza M, Finn MB, Hyman BT, Bacskai BJ, Holtzman DM (2008) Rapida risposta microgliale intorno alla patologia amiloide dopo somministrazione di anticorpi anti-Abeta sistemica nei topi PDAPP. J Neurosci 28 (52): 14156-14164. doi: 10.1523 / jneurosci.4147-08.2008
PubMed PubMedCentral CrossRef Google Scholar
Kool M, Petrilli V, De Smedt T, Rolaz A, Hammad H, van Nimwegen M (2008a) Avanguardia: l'allumino adiuvante stimola le cellule infiammatorie dendritiche attraverso l'attivazione dell'inflammasoma NALP3. J Immunol 181: 3755-3759
PubMed CrossRef Google Scholar
Kool M, Soullie T, van Nimwegen M, Willart MA, Muskens F, Jung S, Hoogsteden HC, Hammad H, Lambrecht BN (2008b) Alum adiuvante aumenta l'immunità adattativa inducendo acido urico e attivando cellule dendritiche infiammatorie. J Exp Med 205 (4): 869-882. doi: 10.1084 / jem.20071087
PubMed PubMedCentral CrossRef Google Scholar
Kool M, Willart MA, van Nimwegen M, Bergen I, Pouliot P, Virchow JC, Rogers N, Osorio F, Reise Sousa C, Hammad H, Lambrecht BN (2011) Un ruolo inaspettato per l'acido urico come induttore di T helper 2 immunità cellulare agli antigeni inalati e mediatore infiammatorio dell'asma allergico. Immunità 34 (4): 527-540. doi: 10.1016 / j.immuni.2011.03.015
PubMed CrossRef Google Scholar
Kool M, Fierens K, Lambrecht BN (2012) Alum adiuvante: alcuni dei trucchi del più antico adiuvante. J Med Microbiol 61 (Pt 7): 927-934. doi: 10.1099 / jmm.0.038943-0
PubMed CrossRef Google Scholar
Korczyn AD, Hassin-Baer S (2015) Il decorso della malattia nella malattia di Parkinson può essere rallentato? BMC Med 13: 295. doi: 10.1186 / s12916-015-0534-x
PubMed PubMedCentral CrossRef Google Scholar
Koufaris C, Sismani C (2015) Modulazione del genoma ed epigenoma di individui suscettibili all'autismo da fattori di rischio ambientale. Int J Mol Sci 16 (4): 8699-8718. doi: 10.3390 / ijms16048699
PubMed PubMedCentral CrossRef Google Scholar
Kumar V, Gill KD (2014) Lo stress ossidativo e la disfunzione mitocondriale nella neurotossicità dell'alluminio e il suo miglioramento: una revisione. Neurotoxicology 41: 154-166. doi: 10.1016 / j.neuro.2014.02.004
PubMed CrossRef Google Scholar
Kumar V, Bal A, Gill KD (2008) Compromissione del metabolismo energetico mitocondriale in diverse regioni del cervello di ratto dopo esposizione cronica all'alluminio. Brain Res 1232: 94-103. doi: 10.1016 / j.brainres.2008.07.028
PubMed CrossRef Google Scholar
Kumar A, Prakash A, Dogra S (2011) Effetto neuroprotettivo del carvedilolo contro la tossicità indotta da alluminio: possibili alterazioni comportamentali e biochimiche nei ratti. Pharmacol Rep 63 (4): 915-923
PubMed CrossRef Google Scholar
Kumar S, Ingle H, Prasad DV, Kumar H (2013) Riconoscimento di infezione batterica da parte di sensori immunitari innati. Crit Rev Microbiol 39 (3): 229-246. doi: 10.3109 / 1040841x.2012.706249
PubMed CrossRef Google Scholar
Kuroda E, Ishii KJ, Uematsu S, Ohata K, Coban C, Akira S, Aritake K, Urade Y, Morimoto Y (2011) I cristalli di silice e i sali di alluminio regolano la produzione di prostaglandina nei macrofagi attraverso i meccanismi indipendenti dall'inflammasoma NALP3. Immunità 34 (4): 514-526. doi: 10.1016 / j.immuni.2011.03.019
PubMed CrossRef Google Scholar
Ladd-Acosta C, Hansen KD, Briem E, Fallin MD, Kaufmann WE, Feinberg AP (2014) Alterazioni di metilazione del DNA comuni in più regioni cerebrali nell'autismo. Mol Psychiatry 19 (8): 862-871. doi: 10.1038 / mp.2013.114
PubMed CrossRef Google Scholar
Lam B, Masellis M, Freedman M, Stuss DT, Black SE (2013) Clinical, imaging e eterogeneità patologica della sindrome di Alzheimer. Alzheimers Res Ther 5 (1): 1. doi: 10.1186 / alzrt155
PubMed PubMedCentral CrossRef Google Scholar
Land WG (2015) Il ruolo dei modelli molecolari associati ai danni nelle malattie umane: parte i - promozione dell'infiammazione e dell'immunità. Sultan Qaboos Univ Med J 15 (1): e9-e21
PubMed PubMedCentral Google Scholar
Landsberg JP, McDonald B, Watt F (1992) Assenza di alluminio nei nuclei della placca neuritica nella malattia di Alzheimer. Nature 360: 65-68
PubMed CrossRef Google Scholar
Vaccini di Langer-Gould A, Qian L, Tartof SY, Brara SM, Jacobsen SJ, Beaber BE, Sy LS, Chao C, Hechter R, Tseng HF (2014) e il rischio di sclerosi multipla e altre malattie demielinizzanti del sistema nervoso centrale. JAMA Neurol 71 (12): 1506-1513. doi: 10.1001 / jamaneurol.2014.2633
PubMed CrossRef Google Scholar
LaSalle JM (2013) Strategie epigenomiche all'interfaccia di fattori di rischio genetici e ambientali per l'autismo. J Hum Genet 58 (7): 396-401. doi: 10.1038 / jhg.2013.49
PubMed PubMedCentral CrossRef Google Scholar
Lee SM, Koh HJ, Park DC, Song BJ, Huh TL, Park JW (2002) Cytosolic NADP (+) - lo stato di isocitrato deidrogenasi dipendente modula il danno ossidativo alle cellule. Gratuito Radic Biol Med 32 (11): 1185-1196
PubMed CrossRef Google Scholar
Lemire J, Appanna VD (2011) Disfunzione di tossicità e astrociti dell'alluminio: un legame metabolico con disturbi neurologici. J Inorg Biochem 105 (11): 1513-1517. doi: 10.1016 / j.jinorgbio.2011.07.001
PubMed CrossRef Google Scholar
Lemire J, Mailloux R, Darwich R, Auger C, Appanna VD (2011) L'interruzione del metabolismo della L-carnitina da parte della tossicità dell'alluminio e dello stress ossidativo promuove la dislipidemia nelle cellule umane astrocitiche ed epatiche. Toxicol Lett 203 (3): 219-226. doi: 10.1016 / j.toxlet.2011.03.019
PubMed CrossRef Google Scholar
Gli aplotipi di Levandowski CB, Mailloux CM, Ferrara TM, Gowan K, Ben S, Jin Y, McFann KK, Holland PJ, Fain PR, Dinarello CA, Spritz RA (2013) NLRP1 associati a vitiligine e autoimmunità aumentano l'elaborazione dell'interleuchina-1beta tramite NLRP1 inflammasome. Proc Natl Acad Sci 110 (8): 2952-2956. doi: 10.1073 / pnas.1222808110
PubMed PubMedCentral CrossRef Google Scholar
Li H, Willingham SB, Ting JP, Re F (2008) Avanguardia: l'attivazione dell'infiammomione da parte dell'allume e l'effetto adiuvante dell'allume sono mediati da NLRP3. J Immunol 181 (1): 17-21
PubMed PubMedCentral CrossRef Google Scholar
Liu Y, Yin H, Zhao M, Lu Q (2014) TLR2 e TLR4 nelle malattie autoimmuni: una revisione completa. Clin Rev Allergy Immunol 47 (2): 136-147. doi: 10.1007 / s12016-013-8402-y
PubMed CrossRef Google Scholar
Llorente-Folch I, Rueda CB, Amigo I, Arco A, Saheki T, Pardo B, Satrustegui J (2013) La regolazione del calcio della respirazione mitocondriale mantiene l'omeostasi dell'ATP e richiede lo shuttle aspartato ARALAR / AGC1-malato nei neuroni corticali intatti. J Neurosci 33 (35): 13957-13971, 13971a. doi: 10.1523 / jneurosci.0929-13.2013
Lovell MA, Ehmann WD, Markesbery WR (1993) Analisi con microsonda laser dell'alluminio del cervello nella malattia di Alzheimer. Ann Neurol 33 (1): 36-42
PubMed CrossRef Google Scholar
Lu H, Hu J, Li J, Pang W, Hu Y, Yang H, Li W, Huang C, Zhang M, Jiang Y (2013) dose ottimale di integrazione di zinco per prevenire neurotossicità indotta da alluminio nei ratti. Neural Regen Res 8 (29): 2754-2762. doi: 10.3969 / j.issn.1673-5374.2013.29.007
PubMed PubMedCentral Google Scholar
Lu X, Liang R, Jia Z, Wang H, Pan B, Zhang Q, Niu Q (2014) Disturbi cognitivi e espressione di tau-proteina tra i lavoratori in fusione di alluminio in pensione. J Occup Environ Med 56 (2): 155-160. doi: 10.1097 / jom.0000000000000100
PubMed CrossRef Google Scholar
Lucas K, Maes M (2013) Ruolo del ciclo radicalico dei recettori Toll Like (TLR) nell'infiammazione cronica: possibili trattamenti mirati alla via del TLR4. Mol Neurobiol 48: 190-204
PubMed CrossRef Google Scholar
Lucas K, Morris G, Anderson G, Maes M (2015) La via del ciclo dei radicali dei recettori del pedaggio: un nuovo target di farmaci nella fatica cronica immuno-correlata. CNS Neurol Disord Drug Targets 14 (7): 838-854
PubMed CrossRef Google Scholar
Lukiw WJ (2012) miRNA proinfiammatori regolati da NF-kappaB nella malattia di Alzheimer. Alzheimers Res Ther 4 (6): 47. doi: 10.1186 / alzrt150
PubMed PubMedCentral CrossRef Google Scholar
Lunnon K, Teeling J, Tutt A, Cragg M, Glennie M, Perry V (2011) L'infiammazione sistemica modula l'espressione del recettore Fc sulla microglia durante la neurodegenerazione cronica. J Immunol 186: 7215-7224
PubMed CrossRef Google Scholar
Ma TY, Tran D, Hoa N, Nguyen D, Merryfield M, Tarnawski A (2000) Meccanismo di regolazione extracellulare del calcio della permeabilità della giunzione stretta epiteliale intestinale: ruolo del coinvolgimento del citoscheletro. Microsc Res Tech 51 (2): 156-168. doi: 10.1002 / 1097-0029 (20001015) 51: 2 <156 :: aid-jemt7> 3.0.co; 2-j
PubMed CrossRef Google Scholar
Mailloux RJ, Appanna VD (2007) La tossicità dell'alluminio innesca la traslocazione nucleare di HIF-1alpha e promuove l'anaerobiosi negli epatociti. Toxicol in Vitro 21 (1): 16-24. doi: 10.1016 / j.tiv.2006.07.013
PubMed CrossRef Google Scholar
Mailloux RJ, Hamel R, Appanna VD (2006) La tossicità dell'alluminio provoca un ciclo di TCA disfunzionale e l'accumulo di succinato negli epatociti. J Biochem Mol Toxicol 20 (4): 198-208. doi: 10.1002 / jbt.201
PubMed CrossRef Google Scholar
Mailloux R, Lemire J, Appanna V (2007) La disfunzione mitocondriale indotta da alluminio porta all'accumulo di lipidi negli epatociti umani: un legame con l'obesità. Cell Physiol Biochem 20 (5): 627-638. doi: 10.1159 / 000107546
PubMed CrossRef Google Scholar
Mailloux RJ, Puiseux-Dao S, Appanna VD (2009) L'alfa-chetoglutarato abroga la localizzazione nucleare di HIF-1alpha in epatociti esposti all'alluminio. Biochimie 91 (3): 408-415. doi: 10.1016 / j.biochi.2008.10.014
PubMed CrossRef Google Scholar
Mailloux RJ, Lemire J, Appanna VD (2011) Risposta epatica alla tossicità dell'alluminio: dislipidemia e malattie del fegato. Exp Cell Res 317 (16): 2231-2238. doi: 10.1016 / j.yexcr.2011.07.009
PubMed CrossRef Google Scholar
Mannello F, Ligi D, Canale M (2013) Alluminio, carbonili e citochine nei fluidi aspirati capezzoli umani: Possibile relazione tra infiammazione, stress ossidativo e microambiente del cancro al seno. J Inorg Biochem 128: 250-256. doi: 10.1016 / j.jinorgbio.2013.07.003
PubMed CrossRef Google Scholar
Marichal T, Ohata K, Bedoret D, Mesnil C, Sabatel C, Kobiyama K, Lekeux P, Coban C, Akira S, Ishii KJ, Bureau F, Desmet CJ (2011) Il DNA rilasciato dalle cellule ospiti morenti media l'attività adiuvante dell'alluminio. Nat Med 17 (8): 996-1002. doi: 10.1038 / nm.2403
PubMed CrossRef Google Scholar
Marrack P, McKee AS, Munks MW (2009) Verso una comprensione dell'azione adiuvante dell'alluminio. Nat Rev Immunol 9 (4): 287-293. doi: 10.1038 / nri2510
PubMed PubMedCentral CrossRef Google Scholar
Marrale M, Albanese NN, Cali F, Romano V (2014) Valutare l'impatto delle varianti del numero di copie sui geni dei miRNA nell'autismo mediante simulazione Monte Carlo. PLoS One 9 (3): e90947. doi: 10.1371 / journal.pone.0090947
PubMed PubMedCentral CrossRef Google Scholar
Marshak-Rothstein A (2006) recettori Toll-like nella malattia sistemica autoimmune. Nat Rev Immunol 6 (11): 823-835
PubMed CrossRef Google Scholar
Matcovitch-Natan O, Winter DR, Giladi A, Vargas Aguilar S, Spinrad A, Sarrazin S, Ben-Yehuda H, David E, Zelada González F, Perrin P, Keren-Shaul H, Gury M, Lara-Astaiso D, Thaiss CA, Cohen M, Bahar Halpern K, Baruch K, Deczkowska A, Lorenzo-Vivas E, Itzkovitz S, Elinav E, Sieweke MH, Schwartz M, Amit I (2016) Lo sviluppo di Microglia segue un programma graduale per regolare l'omeostasi cerebrale. Science 353 (6301): aad8670. doi: 10.1126 / science.aad8670
McDougle CJ, Landino SM, Vahabzadeh A, O'Rourke J, Zurcher NR, Finger BC, Palumbo ML, Helt J, Mullett JE, Hooker JM, Carlezon WA Jr (2015) Verso un sottotipo immuno-mediato di disturbo dello spettro autistico. Brain Res 1617: 72-92. doi: 10.1016 / j.brainres.2014.09.048
PubMed CrossRef Google Scholar
Mead J, Ashwood P (2015) Prove che supportano una risposta immunitaria alterata nell'ASD. Immunol Lett 163 (1): 49-55. doi: 10.1016 / j.imlet.2014.11.006
PubMed CrossRef Google Scholar
Mellios N, Sur M (2012) Il ruolo emergente dei microRNA nella schizofrenia e nei disturbi dello spettro autistico. Front Psychiatry 3:39. doi: 10.3389 / fpsyt.2012.00039
PubMed PubMedCentral CrossRef Google Scholar
Melnyk S, Fuchs G, Schulz E, Lopez M, Kahler S, Fussell J (2012) squilibrio metabolico associato a disregolazione della metilazione e danno ossidativo nei bambini con autismo. J Autism Dev Disord 42: 367-377
PubMed PubMedCentral CrossRef Google Scholar
Michel M, Schmidt MJ, Mirnics K (2012) Disregolazione del gene del sistema immunitario in autismo e schizofrenia. Developl Neurobiol 72 (10): 1277-1287. doi: 10.1002 / dneu.22044
CrossRef Google Scholar
Miranda-Hernandez S, Baxter A (2013) Ruolo dei recettori a pedaggio nella sclerosi multipla. Am J Clin Exp Immunol 2: 75-93
PubMed PubMedCentral Google Scholar
Mohamed Fel B, Zaky EA, El-Sayed AB, Elhossieny RM, Zahra SS, Salah Eldin W, Youssef WY, Khaled RA, Youssef AM (2015) Valutazione di capelli in alluminio, piombo e mercurio in un campione di bambini egiziani autistici: Fattori di rischio ambientale di metalli pesanti nell'autismo. Behav Neurol 2015: 545674. doi: 10.1155 / 2015/545674
PubMed Google Scholar
Mohan N, Alleyne T, Adogwa A (2009) Gli effetti dell'alluminio ingerito sull'attività del citocromo ossidasi cerebrale. West Indian Med J 58 (5): 422-427
PubMed Google Scholar
Molofsky AV, Krenick R, Ullian E, Hh T, Deneen B, Richardson WD, Barres BA, Rowitch DH (2012) Astrociti e malattie: una prospettiva neurosviluppo. Genes Dev 26 (9): 891-907. doi: 10.1101 / gad.188326.112
PubMed PubMedCentral CrossRef Google Scholar
Mor M, Nardone S, Sams DS, Elliott E (2015) Ipometilazione del promotore miR-142 e sovraregolazione dei microRNA che hanno come target il gene del recettore dell'ossitocina nella corteccia prefrontale dell'autismo. Autismo molecolare 6:46. doi: 10.1186 / s13229-015-0040-1
PubMed PubMedCentral CrossRef Google Scholar
Moraes CF, Benedet AL, Souza VC, Lins TC, Camargos EF, Naves JO, Brito CJ, Cordova C, Pereira RW, Nobrega OT (2013) polimorfismi del gene delle citochine e malattia di Alzheimer in Brasile. Neuroimmunomodulazione 20 (5): 239-246. doi: 10.1159 / 000350368
PubMed CrossRef Google Scholar
Morgan J, Chana G, Pardo C, Achim C, Semendeferi K, Buckwalter J (2010) Attivazione microgliale e aumento della densità microgliale osservata nella corteccia prefrontale dorsolaterale nell'autismo. Biol Psychiatry 68: 368-376
PubMed CrossRef Google Scholar
Morgan JT, Chana G, Abramson I, Semendeferi K, Courchesne E, Everall IP (2012) Anormale organizzazione spaziale microglial-neuronale nella corteccia prefrontale dorsolaterale nell'autismo. Brain Res 1456: 72-81. doi: 10.1016 / j.brainres.2012.03.036
PubMed CrossRef Google Scholar
Morris G, Berk M (2015) Le molte strade per la disfunzione mitocondriale nei disturbi neuroimmuni e neuropsichiatrici. BMC Med 13 (1): 68
PubMed PubMedCentral CrossRef Google Scholar
Morris G, Berk M (2016) L'uso putativo del litio nella malattia di Alzheimer. Curr Alzheimer Res 13 (8): 853-861
PubMed CrossRef Google Scholar
Morris G, Maes M (2012) Un modello neuro-immune di encefalomielite mialgica / sindrome da stanchezza cronica. Metab Brain Dis 28: 523-540. doi: 10.1007 / s11011-012-9324-8
Morris G, Maes M (2014) Stress ossidativo e nitrosativo e vie immunitario-infiammatorie in pazienti con Encefalomielite Mialgica (ME) / Sindrome da Stanchezza Cronica (CFS). Curr Neuropharmacol 12: 168-185
PubMed PubMedCentral CrossRef Google Scholar
Morris G, Berk M, Galecki P, Walder K, Maes M (2015a) La patofisiologia neuroimmune dell'affaticamento centrale e periferico nelle malattie sistemico-immuno-infiammatorie e neuroimmuni. Mol Neurobiol 53 (2): 1195-1219. doi: 10.1007 / s12035-015-9090-9
PubMed CrossRef Google Scholar
Morris G, Berk M, Walder K, Maes M (2015b) Vie centrali che causano affaticamento nelle malattie neuroinfiammatorie e autoimmuni. BMC Med 13 (1): 28
PubMed PubMedCentral CrossRef Google Scholar
Mostafa GA, Shehab AA, Al-Ayadhi LY (2013) Il legame tra alcuni alleli sul sistema antigene leucocitario umano e l'autismo nei bambini. J Neuroimmunol 255 (1-2): 70-74. doi: 10.1016 / j.jneuroim.2012.10.002
PubMed CrossRef Google Scholar
Mundalil Vasu M, Anitha A, Thanseem I, Suzuki K, Yamada K, Takahashi T, Wakuda T, Iwata K, Tsujii M, Sugiyama T, Mori N (2014) Profili di siero microRNA in bambini con autismo. Autismo molecolare 5:40. doi: 10.1186 / 2040-2392-5-40
PubMed PubMedCentral CrossRef Google Scholar
Munks MW, McKee AS, Macleod MK, Powell RL, Degen JL, Reisdorph NA, Kappler JW, Marrack P (2010) Gli adiuvanti di alluminio provocano trappole extracellulari fibrinodipendenti in vivo. Blood 116 (24): 5191-5199. doi: 10.1182 / blood-2010-03-275529
PubMed PubMedCentral CrossRef Google Scholar
Murakami K, Yoshino M (2004) L'alluminio diminuisce la rigenerazione del glutatione mediante l'inibizione della NADP-isocitrato deidrogenasi nei mitocondri. J Cell Biochem 93 (6): 1267-1271. doi: 10.1002 / jcb.20261
PubMed CrossRef Google Scholar
Mustafa Rizvi SH, Parveen A, Verma AK, Ahmad I, Arshad M, Mahdi AA (2014) La morte cellulare mediata dallo stress del reticolo endoplasmatico indotto dall'alluminio nella linea cellulare di neuroblastoma SH-SY5Y è indipendente dalla p53. PLoS One 9 (5): e98409. doi: 10.1371 / journal.pone.0098409
PubMed PubMedCentral CrossRef Google Scholar
Nagai T (2013) Attivazione immunitaria innata perinatale e sviluppo neuropsicologico. Nihon Shinkei Seishin Yakurigaku Zasshi 33 (4): 149-154
PubMed Google Scholar
Nampoothiri M, John J, Kumar N, Mudgal J, Nampurath GK, Chamallamudi MR (2015) Ruolo modulatorio della simvastatina contro i cambiamenti comportamentali e biochimici indotti dal cloruro di alluminio nei ratti. Behav Neurol 2015: 210169. doi: 10.1155 / 2015/210169
PubMed PubMedCentral CrossRef Google Scholar
Napoli E, Wong S, Giulivi C (2013) Prove di danno da specie reattive all'ossigeno mediato al DNA mitocondriale nei bambini con autismo tipico. Autismo molecolare 4 (1): 1-8. doi: 10.1186 / 2040-2392-4-2
CrossRef Google Scholar
Nardone S, Sharan Sams D, Reuveni E, Getselter D, Oron O, Karpuj M, Elliott E (2014) l'analisi di metilazione del cervello autistico rivela molteplici percorsi biologici disregolati. Transl Psychiatry 4: e433. doi: 10.1038 / tp.2014.70
PubMed PubMedCentral CrossRef Google Scholar
Netea MG, Wijmenga C, O'Neill LAJ (2012) Variazione genetica dei recettori Toll-like e suscettibilità alla malattia. Nat Immunol 13 (6): 535-542
PubMed CrossRef Google Scholar
Neumann H, Takahashi K (2007) Ruolo essenziale del recettore di attivazione microgliale espresso su cellule mieloidi-2 (TREM2) per l'omeostasi immune del tessuto nervoso centrale. J Neuroimmunol 184 (1-2): 92-99. doi: 10.1016 / j.jneuroim.2006.11.032
PubMed CrossRef Google Scholar
Newairy AS, Salama AF, Hussien HM, Yousef MI (2009) Propoli allevia la perossidazione lipidica indotta dall'alluminio e i parametri biochimici nei ratti maschi. Food Chem Toxicol 47 (6): 1093-1098
PubMed CrossRef Google Scholar
Noriega DB, Savelkoul HF (2014) Disregolazione immunitaria nel disturbo dello spettro autistico. Eur J Pediatr 173 (1): 33-43. doi: 10.1007 / s00431-013-2183-4
PubMed CrossRef Google Scholar
Orihuela D, Meichtry V, Pizarro M (2005) Alterazione indotta dall'alluminio dell'assorbimento transcellulare del calcio nell'intestino tenue: assorbimento del calcio e influenza del glutatione. J Inorg Biochem 99 (9): 1879-1886. doi: 10.1016 / j.jinorgbio.2005.07.003
PubMed CrossRef Google Scholar
Ortiz G, Pacheco-Moises F, Bitzer-Quintero O, Ramirez-Anguiano A, Flores-Alvarado L, Ramirez-Ramirez V (2013) Immunologia e stress ossidativo nella sclerosi multipla: approccio clinico e di base. Clin Dev Immunol 2013: 708659
PubMed PubMedCentral CrossRef Google Scholar
Ottoboni L, Frohlich IY, Lee M, Healy BC, Keenan BT, Xia Z, Chitnis T, Guttmann CR, Khoury SJ, Weiner HL, Hafler DA, De Jager PL (2013) Rilevanza clinica e conseguenze funzionali del locus di sclerosi multipla TNFRSF1A . Neurologia 81 (22): 1891-1899. doi: 10.1212 / 01.wnl.0000436612.66328.8a
PubMed PubMedCentral CrossRef Google Scholar
Pittore MM, Atagi Y, Liu CC, Rademakers R, Xu H, Friggitrice JD, Bu G (2015) TREM2 nell'omeostasi del SNC e malattia neurodegenerativa. Mol Neurodegener 10:43. doi: 10.1186 / s13024-015-0040-9
PubMed PubMedCentral CrossRef Google Scholar
Paradowska-Gorycka A, Jurkowska M (2013) Struttura, modello di espressione e attività biologica del complesso molecolare TREM-2 / DAP12. Hum Immunol 74 (6): 730-737. doi: 10.1016 / j.humimm.2013.02.003
PubMed CrossRef Google Scholar
Paz Soldan MM, Rodriguez M (2002) Eterogeneità della patogenesi nella sclerosi multipla: implicazioni per la promozione della reielinizzazione. Journal of Infectious Diseases 186 (Supplemento 2): S248-S253. doi: 10,1086 / 344283
Pellegrino P, Clementi E, Radice S (2015) sugli adiuvanti e l'autoimmunità del vaccino: prove attuali e prospettive future. Autoimmun Rev 14 (10): 880-888. doi: 10.1016 / j.autrev.2015.05.014
PubMed CrossRef Google Scholar
Pelly L, Vardy C, Fernandez B, Newhook LA, Chafe R (2015) Incidenza e prevalenza di coorte per disturbi dello spettro autistico nella penisola di Avalon, Terranova e Labrador. CMAJ Open 3 (3): E276-E280. doi: 10.9778 / cmajo.20140056
PubMed PubMedCentral CrossRef Google Scholar
Peng Q, Long CL, Malhotra S, Humphrey MB (2013) È necessaria un'interazione fisica tra le proteine dell'adattatore DOK3 e DAP12 per inibire la segnalazione del lipopolisaccaride nei macrofagi. Sci Signal 6 (289): ra72. doi: 10.1126 / scisignal.2003801
PubMed PubMedCentral CrossRef Google Scholar
Perricone C, Colafrancesco S, Mazor RD, Soriano A, Agmon-Levin N, Shoenfeld Y (2013) Sindrome autoimmune / infiammatoria indotta da adiuvanti (ASIA) 2013: Svelare gli aspetti patogenetici, clinici e diagnostici. J Autoimmun 47: 1-16. doi: 10.1016 / j.jaut.2013.10.004
PubMed CrossRef Google Scholar
Perry V, Holmes C (2014) Microglial priming nella malattia neurodegenerativa. Nat Rev Neurol 10: 217-224
PubMed CrossRef Google Scholar
Perry V, Nicoll J, Holmes C (2010) Microglia nella malattia neurodegenerativa. Nat Rev Neurol 6: 193-201
PubMed CrossRef Google Scholar
Petrovsky N (2015) Sicurezza comparativa degli adiuvanti vaccinali: un riassunto delle prove attuali e delle esigenze future. Drug Saf 38 (11): 1059-1074. doi: 10.1007 / s40264-015-0350-4
PubMed PubMedCentral CrossRef Google Scholar
Piccinini AM, Midwood KS (2010) DAMPANTE l'infiammazione modulando la segnalazione TLR. Mediat Inflamm 2010. doi: 10.1155 / 2010/672395
Pineton de Chambrun G, Body-Malapel M, Frey-Wagner I, Djouina M, Deknuydt F, Atrott K, Esquerre N, Altare F, Neut C, Arrieta MC, Kanneganti TD, Rogler G, Colombel JF, Cortot A, Desreumaux P , Vignal C (2014) L'alluminio aumenta l'infiammazione e diminuisce la guarigione della mucosa nella colite sperimentale nei topi. Mucosal Immunol 7 (3): 589-601. doi: 10.1038 / mi.2013.78
PubMed CrossRef Google Scholar
Pogue AI, Li YY, Cui JG, Zhao Y, Kruck TP, Percy ME, Tarr MA, Lukiw WJ (2009) Caratterizzazione di un down-regulation mediato da miRNA-146a del fattore di complemento H (CFH) regolato da NF-kappaB in cellule cerebrali umane stressate dal metallo-solfato. J Inorg Biochem 103 (11): 1591-1595. doi: 10.1016 / j.jinorgbio.2009.05.012
PubMed CrossRef Google Scholar
Polonia GA, Kennedy RB, McKinney BA, Ovsyannikova IG, Lambert ND, Jacobson RM, Oberg AL (2013) Vaccinomica, adversomica e teoria della rete di risposta immunitaria: vaccinologia individualizzata nel 21 ° secolo. Semin Immunol 25 (2): 89-103. doi: 10.1016 / j.smim.2013.04.007
PubMed PubMedCentral CrossRef Google Scholar
Poliani PL, Wang Y, Fontana E, Robinette ML, Yamanishi Y, Gilfillan S, Colonna M (2015) TREM2 sostiene l'espansione microgliale durante l'invecchiamento e la risposta alla demielinizzazione. J Clin Invest 125 (5): 2161-2170. doi: 10.1172 / jci77983
PubMed PubMedCentral CrossRef Google Scholar
Pontillo A, Girardelli M, Kamada AJ, Pancotto JA, Donadi EA, Crovella S, Sandrin-Garcia P (2012) I polimorfismi nei geni inflammasoma sono coinvolti nella predisposizione al lupus eritematoso sistemico. Autoimmunità 45 (4): 271-278. doi: 10.3109 / 08916934.2011.637532
PubMed CrossRef Google Scholar
Powell BS, Andrianov AK, Fusco PC (2015) Coadiuvanti del vaccino polionico: un altro sguardo ai sali di alluminio e ai polielettroliti. Clin Exper Vacc Res 4 (1): 23-45. doi: 10.7774 / cevr.2015.4.1.23
CrossRef Google Scholar
Pradhan VD, Das S, Surve P, Ghosh K (2012) Recettori toll-like in autoimmunità con particolare riferimento al lupus eritematoso sistemico. Indian J Human Gene 18 (2): 155-160. doi: 10.4103 / 0971-6866.100750
CrossRef Google Scholar
Pramparo T, Pierce K, Lombardo MV, Carter Barnes C, Marinero S, Ahrens-Barbeau C, Murray SS, Lopez L, Xu R, Courchesne E (2015) Pronostico dell'autismo mediante traduzione e geni congeniti / infiammatori espressi nei bambini da pediatria pratiche comunitarie. JAMA Psychiatry 72 (4): 386-394. doi: 10.1001 / jamapsychiatry.2014.3008
PubMed CrossRef Google Scholar
Rainbolt TK, Saunders JM, Wiseman RL (2014) Regolazione reattiva dello stress dei mitocondri attraverso la risposta proteica dispiegata ER. Tendenze Endocrinol Metab 25 (10): 528-537. doi: 10.1016 / j.tem.2014.06.007
PubMed CrossRef Google Scholar
Ramos PS, Sajuthi S, Langefeld CD, Walker SJ (2012) I geni della funzione immunitaria CD99L2, JARID2 e TPO mostrano associazione con il disturbo dello spettro autistico. Autismo molecolare 3: 4-4. doi: 10.1186 / 2040-2392-3-4
PubMed PubMedCentral CrossRef Google Scholar
Rangasamy S, D'Mello SR, Narayanan V (2013) Epigenetica, spettro autistico e disturbi del neurosviluppo. Neurotherapeutics 10 (4): 742-756. doi: 10.1007 / s13311-013-0227-0
PubMed PubMedCentral CrossRef Google Scholar
Rapoport JL, Giedd JN, Gogtay N (2012) Modello neurodevelopmental della schizofrenia: aggiornamento 2012. Mol Psychiatry 17 (12): 1228-1238. doi: 10.1038 / mp.2012.23
PubMed PubMedCentral CrossRef Google Scholar
Reuter S, Gupta S, Chaturvedi M, Aggarwal B (2010) Stress ossidativo, infiammazione e cancro: come sono collegati? Gratuito Radic Biol Med 49: 1603-1616
PubMed PubMedCentral CrossRef Google Scholar
Roberts AL, Lyall K, Hart JE, Laden F, Solo AC, Bobb JF, Koenen KC, Ascherio A, Weisskopf MG (2013) Esposizione agli inquinanti atmosferici perinatali e disturbo dello spettro autistico nei bambini dei partecipanti allo Studio di salute degli infermieri II. Environ Health Perspect 121 (8): 978-984. doi: 10.1289 / ehp.1206187
PubMed PubMedCentral Google Scholar
Rondeau V, Jacqmin-Gadda H, Commendes D, Helmer C, Dartigues JF (2009) Alluminio e silice nell'acqua potabile e il rischio di malattia di Alzheimer o declino cognitivo: risultati del follow-up a 15 anni della coorte PAQUID. Am J Epidemiol 169 (4): 489-496. doi: 10.1093 / aje / kwn348
PubMed CrossRef Google Scholar
Rossignol D, Frye R (2014) Evidenza che collega lo stress ossidativo, la disfunzione mitocondriale e l'infiammazione nel cervello di individui con autismo. Front Physiol 5: 150
PubMed PubMedCentral CrossRef Google Scholar
Rueda CB, Llorente-Folch I, Amigo I, Contreras L, Gonzalez-Sanchez P, Martinez-Valero P, Juaristi I, Pardo B, Arco A, Satrustegui J (2014) Ca (2+) regolazione della funzione mitocondriale nei neuroni . Biochim Biophys Acta 1837 (10): 1617-1624. doi: 10.1016 / j.bbabio.2014.04.010
PubMed CrossRef Google Scholar
Ruggeri B, Sarkans U, Schumann G, Persico AM (2014) Biomarcatori nel disturbo dello spettro autistico: il vecchio e il nuovo. Psychopharmacology 231 (6): 1201-1216. doi: 10.1007 / s00213-013-3290-7
PubMed CrossRef Google Scholar
Rutter M (2005) Incidenza dei disturbi dello spettro autistico: cambiamenti nel tempo e il loro significato. Acta Paediatr 94 (1): 2-15
PubMed CrossRef Google Scholar
Sahoo T, Theisen A, Rosenfeld JA, Lamb AN, Ravnan JB, Schultz RA, Torchia BS, Neill N, Casci I, Bejjani BA, Shaffer LG (2011) Copia varianti di numero di locus di suscettibilità alla schizofrenia sono associati a uno spettro di parole e ritardi di sviluppo e problemi comportamentali. Gene Med 13 (10): 868-880. doi: 10.1097 / GIM.0b013e3182217a06
CrossRef Google Scholar
Saresella M, Piancone F, Marventano I, Zoppis M, Hernis A, Zanette M, Trabattoni D, Chiappedi M, Ghezzo A, Canevini MP, la Rosa F, Esposito S, Clerici M (2016) I complessi di inflammasoma multipli sono attivati nello spettro autistico disturbi. Brain Behav Immun 57: 125-133. doi: 10.1016 / j.bbi.2016.03.009
PubMed CrossRef Google Scholar
Scheller NM, Svanstrom H, Pasternak B, Arnheim-Dahlstrom L, Sundstrom K, Fink K, Hviid A (2015) vaccinazione quadrivalente HPV e rischio di sclerosi multipla e altre malattie demielinizzanti del sistema nervoso centrale. JAMA 313 (1): 54-61. doi: 10.1001 / jama.2014.16946
PubMed CrossRef Google Scholar
Schepens MA, Schonewille AJ, Vink C, van Schothorst EM, Kramer E, Hendriks T, Brummer RJ, Keijer J, van der Meer R, Bovee-Oudenhoven IM (2009) Il calcio supplementare attenua l'aumento correlato alla colite della diarrea, permeabilità intestinale e rotture della matrice extracellulare in ratti transgenici HLA-B27. J Nutr 139 (8): 1525-1533. doi: 10.3945 / jn.109.105205
PubMed CrossRef Google Scholar
Sharma DR, Sunkaria A, Wani WY, Sharma RK, Kandimalla RJL, Bal A, Gill KD (2013) Lo stress ossidativo indotto dall'alluminio determina una diminuzione della biogenesi mitocondriale tramite modulazione dell'espressione di PGC-1α. Toxicol Appl Pharmacol 273 (2): 365-380. doi: 10.1016 / j.taap.2013.09.012
PubMed CrossRef Google Scholar
Sharp FA, Ruane D, Claass B, Creagh E, Harris J, Malyala P, Singh M, O'Hagan DT, Petrilli V, Tschopp J, O'Neill LA, Lavelle EC (2009) Assorbimento di coadiuvanti del vaccino particolato da parte delle cellule dendritiche attiva l'inflammasoma NALP3. Proc Natl Acad Sci USA 106 (3): 870-875. doi: 10.1073 / pnas.0804897106
PubMed PubMedCentral CrossRef Google Scholar
Shastri A, Bonifati DM, Kishore U (2013) Immunità innata e Neuroinfiammazione. Mediat Inflamm 2013: 342931. doi: 10.1155 / 2013/342931
CrossRef Google Scholar
Shaw CA, Tomljenovic L (2013) Alluminio nel sistema nervoso centrale (SNC): tossicità negli esseri umani e negli animali, coadiuvanti del vaccino e autoimmunità. Immunol Res 56 (2-3): 304-316. doi: 10.1007 / s12026-013-8403-1
PubMed CrossRef Google Scholar
Shi Y, Evans JE, Rock KL (2003) Identificazione molecolare di un segnale di pericolo che allerta il sistema immunitario alle cellule morenti. Nature 425 (6957): 516-521. doi: 10.1038 / nature01991
PubMed CrossRef Google Scholar
Shoenfeld Y, Agmon-Levin N (2011) 'ASIA' - sindrome autoimmune / infiammatoria indotta da adiuvanti. J Autoimmun 36 (1): 4-8. doi: 10.1016 / j.jaut.2010.07.003
PubMed CrossRef Google Scholar
Singhal G, Jaehne EJ, Corrigan F, Toben C, Baune BT (2014) Inflammasomes in neuroinfiammazione e cambiamenti nella funzione cerebrale: una revisione focalizzata. Front Neurosci 8 (315). doi: 10.3389 / fnins.2014.00315
Siniscalco D (2015) Commento: L'impatto delle alterazioni neuroimmuni nel disturbo dello spettro autistico. Front Psychiatry 6: 145. doi: 10.3389 / fpsyt.2015.00145
PubMed PubMedCentral CrossRef Google Scholar
Siniscalco D, Cirillo A, Bradstreet JJ, Antonucci N (2013) Reperti epigenetici nell'autismo: nuove prospettive per la terapia. Int J Environ Res Public Health 10 (9): 4261-4273. doi: 10.3390 / ijerph10094261
PubMed PubMedCentral CrossRef Google Scholar
Sloan SA, Barres BA (2013) Glia come driver principali della neuropatologia nelle proteinopatie TDP-43. Proc Natl Acad Sci USA 110 (12): 4439-4440. doi: 10.1073 / pnas.1301608110
PubMed PubMedCentral CrossRef Google Scholar
Smeeth L, Cook C, Fombonne E, Heavey L, Rodrigues LC, Smith PG, Hall AJ (2004) vaccinazione MMR e disturbi pervasivi dello sviluppo: uno studio caso-controllo. Lancet 364 (9438): 963-969. doi: 10.1016 / s0140-6736 (04) 17020-7
PubMed CrossRef Google Scholar
Soriano A, Nesher G, Shoenfeld Y (2015) Previsione dell'autoimmunità post-vaccinazione: chi potrebbe essere a rischio? Pharmacol Res 92: 18-22. doi: 10.1016 / jrsrs.2014.08.002
PubMed CrossRef Google Scholar
Stetson DB, Medzhitov R (2006) Il riconoscimento del DNA citosolico attiva una risposta immunitaria innata dipendente da IRF3. Immunità 24 (1): 93-103. doi: 10.1016 / j.immuni.2005.12.003
PubMed CrossRef Google Scholar
Struys-Ponsar C, Guillard O, van den Bosch de Aguilar P (2000) Effetti dell'esposizione dell'alluminio sul metabolismo del glutammato: una possibile spiegazione della sua tossicità. Exp Neurol 163 (1): 157-164. doi: 10.1006 / exnr.2000.7355
PubMed CrossRef Google Scholar
Su X, Federoff H (2014) Risposte immunitarie nella malattia di Parkinson: interazione tra sistema immunitario centrale e periferico. Biomed Res Int 2014: 275178
PubMed PubMedCentral Google Scholar
Suarez-Fernandez MB, Soldado AB, Sanz-Medel A, Vega JA, Novelli A, Fernandez-Sanchez MT (1999) La degenerazione degli astrociti indotta dall'alluminio avviene attraverso l'apoptosi e provoca la morte neuronale. Brain Res 835 (2): 125-136
PubMed CrossRef Google Scholar
Sumathi T, Shobana C, Thangarajeswari M, Usha R (2015) Effetto protettivo della L-teanina contro neurotossicità indotta dall'alluminio nella corteccia cerebrale, nell'ippocampo e nel cervelletto dell'approccio cerebrale - istopatologico e biochimico del ratto. Drug Chem Toxicol 38 (1): 22-31. doi: 10.3109 / 01480545.2014.900068
PubMed CrossRef Google Scholar
Suzuki K, Sugihara G, Ouchi Y, Nakamura K, Futatsubashi M, Takebayashi K, Yoshihara Y, Omata K, Matsumoto K, Tsuchiya KJ, Iwata Y, Tsujii M, Sugiyama T, Mori N (2013) Attivazione microgliale in giovani adulti con disturbo dello spettro autistico. JAMA Psychiatry 70 (1): 49-58. doi: 10.1001 / jamapsychiatry.2013.272
PubMed CrossRef Google Scholar
Takahashi K, Rochford CD, Neumann H (2005) Clearance dei neuroni apoptotici senza infiammazione da parte del recettore di innesco microgliale espresso su cellule mieloidi-2. J Exp Med 201 (4): 647-657. doi: 10.1084 / jem.20041611
PubMed PubMedCentral CrossRef Google Scholar
Takahashi K, Prinz M, Stagi M, Chechneva O, Neumann H (2007) I precursori mieloidi trasdotti TREM2 mediano la clearance dei detriti tissutali nervosi e facilitano il recupero in un modello animale di sclerosi multipla. PLoS Med 4 (4): e124. doi: 10.1371 / journal.pmed.0040124
PubMed PubMedCentral CrossRef Google Scholar
Tammimies K, Marshall CR, Walker S, Kaur G, Thiruvahindrapuram B, Lionel AC, Yuen RK, Uddin M, Roberts W, Weksberg R, Woodbury-Smith M, Zwaigenbaum L, Anagnostou E, Wang Z, Wei J, Howe JL, Gazzellone MJ, Lau L, Sung WW, Whitten K, Vardy C, Crosbie V, Tsang B, D'Abate L, Tong WW, Luscombe S, Doyle T, Carter MT, Szatmari P, Stuckless S, Merico D, Stavropoulos DJ, Scherer SW, Fernandez BA (2015) Rendimento diagnostico molecolare dell'analisi microarray cromosomica e sequenziamento dell'intero esoma in bambini con disturbo dello spettro autistico. JAMA 314 (9): 895-903. doi: 10.1001 / jama.2015.10078
PubMed CrossRef Google Scholar
I polimorfismi Tan MS, Yu JT, Jiang T, Zhu XC, Wang HF, Zhang W, Wang YL, Jiang W, Tan L (2013) NLRP3 sono associati alla malattia di Alzheimer ad esordio tardivo in cinese Han. J Neuroimmunol 265 (1-2): 91-95. doi: 10.1016 / j.jneuroim.2013.10.002
PubMed CrossRef Google Scholar
Taylor LE, Swerdfeger AL, Eslick GD (2014) I vaccini non sono associati all'autismo: una meta-analisi basata sull'evidenza di studi caso-controllo e di coorte. Vaccino 32 (29): 3623-3629. doi: 10.1016 / j.vaccine.2014.04.085
PubMed CrossRef Google Scholar
Thompson MR, Kaminski JJ, Kurt-Jones EA, Fitzgerald KA (2011) recettori di riconoscimento dei pattern e risposta immunitaria innata all'infezione virale. Virus 3 (6): 920-940. doi: 10.3390 / v3060920
PubMed PubMedCentral CrossRef Google Scholar
Thrash JC, Torbett BE, Carson MJ (2009) Regolazione dello sviluppo dell'espressione TREM2 e DAP12 nel CNS murino: implicazioni per la malattia di Nasu-Hakola. Neurochem Res 34 (1): 38-45. doi: 10.1007 / s11064-008-9657-1
PubMed CrossRef Google Scholar
Tordjman S, Somogyi E, Coulon N, Kermarrec S, Cohen D, Bronsard G, Bonnot O, Weismann-Arcache C, Botbol M, Lauth B, Ginchat V, Roubertoux P, Barburoth M, Kovess V, Geoffray MM, Xavier J ( 2014) Interazioni Gene x Ambiente nei disturbi dello spettro autistico: ruolo dei meccanismi epigenetici. Front Psychiatry 5:53. doi: 10.3389 / fpsyt.2014.00053
PubMed PubMedCentral CrossRef Google Scholar
Torres AR, Westover JB, Rosenspire AJ (2012) HLA Immune Function Genes in Autism. Autism Res Treat 2012: 959073. doi: 10.1155 / 2012/959073
PubMed PubMedCentral Google Scholar
Tunçbilek S (2014) Relazione tra polimorfismi del gene delle citochine e infezione da virus dell'epatite B cronica. World J Gastroenterol 20 (20): 6226-6235. doi: 10.3748 / wjg.v20.i20.6226
PubMed PubMedCentral CrossRef Google Scholar
Vaishnavi V, Manikandan M, Tiwary BK, Munirajan AK (2013) Approfondimenti sull'impatto funzionale dei microRNA presenti nelle varianti del numero di copie associate all'autismo. PLoS One 8 (2): e56781. doi: 10.1371 / journal.pone.0056781
PubMed PubMedCentral CrossRef Google Scholar
Vannuvel K, Renard P, Raes M, Arnould T (2013) Impatto funzionale e morfologico dello stress da ER sui mitocondri. J Cell Physiol 228 (9): 1802-1818. doi: 10.1002 / jcp.24360
PubMed CrossRef Google Scholar
Venegas C, Heneka MT (2017) Modelli molecolari associati ai pericoli nella malattia di Alzheimer. J Leukoc Biol 101 (1): 87-98. doi: 10.1189 / jlb.3MR0416-204R
PubMed CrossRef Google Scholar
Vera-Lastra O, Medina G, Cruz-Dominguez M, Jara L, Shoenfeld Y (2013) Sindrome autoimmune / infiammatoria indotta dagli adiuvanti (sindrome di Shoenfeld): spettro clinico e immunologico. Expert Rev Clin Immunol 9: 361-373
PubMed CrossRef Google Scholar
Verstraeten T, Davis RL, DeStefano F, Lieu TA, Rhodes PH, Black SB, Shinefield H, Chen RT (2003) Sicurezza dei vaccini contenenti timerosal: uno studio in due fasi dei database di organizzazioni di manutenzione sanitaria computerizzate. Pediatrics 112 (5): 1039-1048
PubMed Google Scholar
Voineagu I (2012) Studi sull'espressione genica nell'autismo: passaggio dal genoma al transcriptoma e oltre. Neurobiol Dis 45 (1): 69-75. doi: 10.1016 / j.nbd.2011.07.017
PubMed CrossRef Google Scholar
Voineagu I, Eapen V (2013) Percorsi convergenti nei disturbi dello spettro autistico: interazione tra disfunzione sinaptica e risposte immunitarie. Front Hum Neurosci 7: 738. doi: 10.3389 / fnhum.2013.00738
PubMed PubMedCentral CrossRef Google Scholar
Voineagu I, Wang X, Johnston P, Lowe J, Tian Y, Horvath S (2011) L'analisi trascrittomica del cervello autistico rivela una patologia molecolare convergente. Nature 474: 380-384
PubMed PubMedCentral CrossRef Google Scholar
Walton JR (2006) Alluminio in neuroni ippocampali da umani con malattia di Alzheimer. Neurotoxicology 27 (3): 385-394
PubMed CrossRef Google Scholar
Walton JR (2012) L'interruzione dell'alluminio dell'omeostasi del calcio e la trasduzione del segnale assomigliano al cambiamento che si verifica nell'invecchiamento e nella malattia di Alzheimer. J Alzheimers Dis 29 (2): 255-273. doi: 10.3233 / jad-2011-111712
PubMed Google Scholar
Walton JR (2014) L'assunzione cronica di alluminio causa la malattia di Alzheimer: applicazione dei criteri di causalità di Sir Austin Bradford Hill. J Alzheimers Dis 40 (4): 765-838. doi: 10.3233 / jad-132204
PubMed Google Scholar
Wang Y, Cella M, Mallinson K, Ulrich JD, Young KL, Robinette ML, Gilfillan S, Krishnan GM, Sudhakar S, Zinselmeyer BH, Holtzman DM, Cirrito JR, Colonna M (2015) TREM2 lipid sensing sostiene la risposta microgliale in un Modello di malattia di Alzheimer. Cella 160 (6): 1061-1071. doi: 10.1016 / j.cell.2015.01.049
PubMed PubMedCentral CrossRef Google Scholar
Wang Z, Wei X, Yang J, Suo J, Chen J, Liu X, Zhao X (2016) Esposizione cronica all'alluminio e rischio di malattia di Alzheimer: una meta-analisi. Neurosci Lett 610: 200-206. doi: 10.1016 / j.neulet.2015.11.014
PubMed CrossRef Google Scholar
Willhite CC, Karyakina NA, Yokel RA, Yenugadhati N, Wisniewski TM, Arnold IM, Momoli F, Krewski D (2014) Revisione sistematica dei potenziali rischi per la salute derivanti da esposizioni farmaceutiche, occupazionali e dei consumatori su alluminio metallico e nanoscala, ossidi di alluminio, alluminio idrossido e suoi sali solubili. Crit Rev Toxicol 44 (Suppl 4): 1-80. doi: 10.3109 / 10408444.2014.934439
PubMed PubMedCentral CrossRef Google Scholar
Wong C, Meaburn E, Ronald A, Price T, Jeffries A, Schalkwyk L (2014) Analisi metilomica di gemelli monozigoti discordanti per disturbo dello spettro autistico e relativi tratti comportamentali. Mol Psychiatry 19: 495-503
PubMed CrossRef Google Scholar
Yang CA, Chiang BL (2015) Inflammasomes e autoimmunità umana: una revisione completa. J Autoimmun 61: 1-8. doi: 10.1016 / j.jaut.2015.05.001
PubMed CrossRef Google Scholar
Yang Y, Higashimori H, Morel L (2013) Sviluppo evolutivo degli astrociti e patogenesi dei disturbi dello sviluppo neurologico. J Neurodev Disord 5 (1): 22. doi: 10.1186 / 1866-1955-5-22
PubMed PubMedCentral CrossRef Google Scholar
Yasuda H, Tsutsui T (2013) Valutazione degli squilibri minerali infantili nei disturbi dello spettro autistico (ASD). Int J Environ Res Public Health 10 (11): 6027-6043. doi: 10.3390 / ijerph10116027
PubMed PubMedCentral CrossRef Google Scholar
Yasui M, Kihira T, Ota K (1992) Concentrazioni di calcio, magnesio e alluminio nella malattia di Parkinson. Neurotoxicology 13 (3): 593-600
PubMed Google Scholar
Yousef MI (2004) Cambiamenti indotti dall'alluminio nei parametri emato-biochimici, perossidazione lipidica e attività enzimatica dei conigli maschi: ruolo protettivo dell'acido ascorbico. Tossicologia 199 (1): 47-57. doi: 10.1016 / j.tox.2004.02.014
PubMed CrossRef Google Scholar
Yuan CY, Lee YJ, Hsu GS (2012) Il sovraccarico di alluminio aumenta lo stress ossidativo in quattro aree funzionali del cervello di ratti neonatali. J Biomed Sci 19:51. doi: 10.1186 / 1423-0127-19-51
PubMed PubMedCentral CrossRef Google Scholar
Yumoto S, Kakimi S, Ohsaki A, Ishikawa A (2009) Dimostrazione dell'alluminio nelle fibre amiloidi nei nuclei delle placche senili nel cervello dei pazienti con malattia di Alzheimer. J Inorg Biochem 103 (11): 1579-1584. doi: 10.1016 / j.jinorgbio.2009.07.023
PubMed CrossRef Google Scholar
Zafrir Y, Agmon-Levin N, Paz Z, Shilton T, Shoenfeld Y (2012) Autoimmunità dopo il vaccino contro l'epatite B come parte dello spettro della "Sindrome Autoimmune (Autoinfiammatoria) indotta da adiuvanti" (ASIA): analisi di 93 casi . Lupus 21 (2): 146-152. doi: 10.1177 / 0961203311429318
PubMed CrossRef Google Scholar
Zaky A, Mohammad B, Moftah M, Kandeel K, Bassiouny A (2013) Apurinic / apyrimidinic endonuclease 1 è un modulatore chiave della neuroinfiammazione indotta da alluminio. BMC Neurosci 14 (1): 26
PubMed PubMedCentral CrossRef Google Scholar
Zhang YW, Thompson R, Zhang H, Xu H (2011) elaborazione APP nella malattia di Alzheimer. Mol Brain 4: 3. doi: 10.1186 / 1756-6606-4-3
PubMed PubMedCentral CrossRef Google Scholar
Zhang Y, Yamamoto T, Hisatome I, Li Y, Cheng W, Sole N, Cai B, Huang T, Zhu Y, Li Z, Jing X, Zhou R, Cheng J (2013) L'acido urico induce lo stress ossidativo e l'inibizione della crescita di attivazione di chinasi di proteina attivata da adenosina monofosfato e vie segnale segnale chinasi extracellulare segnale-regolate in cellule β pancreatiche. Mol Cell Endocrinol 375 (1-2): 89-96. doi: 10.1016 / j.mce.2013.04.027
PubMed CrossRef Google Scholar
Zhao Y, Lukiw WJ (2013) segnalazione TREM2, miRNA-34a e l'estinzione della fagocitosi. Front Cell Neurosci 7: 131. doi: 10.3389 / fncel.2013.00131
PubMed PubMedCentral Google Scholar
Zhao Y, Bhattacharjee S, Jones BM, Dua P, Alexandrov PN, Hill JM, Lukiw WJ (2013) Regolazione dell'espressione TREM2 da parte di un NF-piccolo ka, miRNA-34a Cyrillic-sensitive. Neuroreport 24 (6): 318-323. doi: 10.1097 / WNR.0b013e32835fb6b0
PubMed PubMedCentral CrossRef Google Scholar
Zhao Y, Bhattacharjee S, Jones BM, Hill J, Dua P, Lukiw WJ (2014) Regolazione della segnalazione neurotropica da parte del miRNA-125b inducibile, sensibile a NF-kb nella malattia di Alzheimer (AD) e nella glia neuronale umana primaria ( Cellule HNG). Mol Neurobiol 50 (1): 97-106. doi: 10.1007 / s12035-013-8595-3
PubMed CrossRef Google Scholar
Zhubi A, Cook EH, Guidotti A, Grayson DR (2014) Meccanismi epigenetici nel disturbo dello spettro autistico. Int Rev Neurobiol 115: 203-244. doi: 10.1016 / b978-0-12-801311-3.00006-8
PubMed CrossRef Google Scholar
Zielke HR, Jackson MJ, Tildon JT, Max SR (1993) Un meccanismo glutammatergico per la tossicità dell'alluminio negli astrociti. Mol Chem Neuropathol 19 (3): 219-233
PubMed CrossRef Google Scholar












